1. Ambush predators: Pitvipers around the world
Venom is a remarkable biological trait that has evolved independently in numerous organisms on our planet. However, this trait, which characterizes some groups of snakes, culturally generates mixed reactions and attitudes, evoking fear, fascination, and respect at the same time, making snakes stand out among all species in the animal kingdom as creatures of power
[1]. Nevertheless, in the entire world only a handful of snake species exhibit exceptional venom potency or pose a significant threat owing to their sophisticated venom delivery mechanisms and specialized venom composition (see Chapter 5). Approximately 10% of all living snakes belong to the Viperidae family, encompassing vipers, adders, and pitvipers
[2,3]. This family is phylogenetically divided into three subfamilies: Azemiopinae (Fea vipers), Crotalinae (pitvipers), and Viperinae (“true vipers” or “vipers without pits”).
Currently, about 383 species of viperids are recognized, grouped into 37 genera
[4]. Thirteen of these genera are distributed across America:
Agkistrodon[5],
Atropoides[6],
Bothriechis[7],
Bothrocophias[8],
Bothrops[9],
Cerrophidion[10],
Crotalus[11],
Lachesis[12],
Metlapilcoatlus[13],
Mixcoatlus[14],
Ophryacus[15],
Porthidium[16],
Sistrurus[17]; and 24 genera are distributed among the Asian, European and African continents:
Atheris[18],
Azemiops[19],
Bitis[20],
Calloselasma[21],
Causus[22],
Cerastes[11],
Craspedocephalus
[23],
Daboia[20],
Deinagkistrodon[24],
Echis[25],
Eristicophis[26],
Garthius[27],
Gloydius[28],
Hypnale[29],
Macropvipera[30],
Montatheris[31],
Montivipera[32],
Ovophis[33],
Proatheris[31],
Protobothrops[34],
Pseudoceraste s
[35],
Trimeresurus[36],
Tropidolaemus[22], and
Vipera[37].
Among all venomous snakes, the highly specialized venom delivery system of viperids is characterized by tubular front fangs enclosed in venom-conducting canals positioned on a mobile maxillary bone
[38] (see Chapter 5). The specialized long teeth developed by viperids represent one of the most remarkable functional traits linked to their life history. They are primarily sedentary ambush predators that consume mostly hot-blood prey during their adulthood
[38,39]. Consequently, the efficacy of this sophisticated venom release mechanism in hunting, defense, and deterrent functions has resulted in the designation of several viper genera as medically important due to the elevated annual incidence of snakebite (see Chapter 9).
In these ambush hunters, venom composition limits prey selection; hence, feeding has notorious adaptations and specificities. For example, newborn or young snakes exhibit greater lethal activity than those of their adult counterparts
[40], indicating that these biochemical differences are linked with the diet changes and the evolution of the prey. Indeed, proteomic studies have revealed that ontogenetic variation in venom composition is a significant indicator for understanding toxin dynamics and actions for the two most significant Colombian pitvipers:
Bothrops atrox[41] and
B. asper[42,43].
To understand the evolution of snakes from a mechanical means of dominating prey (constriction) to a chemical means (venom injection
[44]), it is imperative to first comprehend the many processes through which natural selection has favored the harmony between divergence and adaptation. This balance has allowed the emergence of vast heterogeneity in the composition, function, and action of snake venom toxins. This chapter provides a brief overview of the past 50 years of research on the evolutionary and natural history of pitviper species that inhabit the tropical ecosystems of Colombia, focusing on the medically important snake species.
2. The evolutionary and geographic perspectives of pitviper colonization and diversification in the South American tropics
One of the great and pioneering contributions in the systematic and biology of pitvipers corresponds to the work of William L. Burger
[45], who clarified the delimitation of several groups and the description of new characters of taxonomic use for the Viperidae family. His pioneering work suggested the division of the large
Bothrops group into five morphologically divergent genera:
Bothriechis, Bothriopsis, Bothrops, Ophryacus and
Porthidium. Nowadays, the validity of
Bothriopsis as a separate genus of snakes is contentious. Historically,
Bothriopsis was considered a valid genus, but phylogenetic and genetic studies have questioned its distinction from the genus
Bothrops. In fact, Burger’s work was crucial because it triggered a series of rearrangements that allowed the description of genera derived from
Porthidium, such as
Atropoides[6] and
Cerrophidion
[10].
Later, Avise was one of the pioneers
[46] to examine hypotheses of phylogenetic and phylogeographic inference in pitvipers from a genetic perspective used molecular markers. He studied the ancestral relationships that are unaffected by convergent selection in similar natural history traits by assessing molecular markers as independent evidence, assuming that these traits were unlikely to be influenced by the same selective pressures that act on morphological traits
[47]. Since the 1990s, DNA sequences have been used to calculate the historical timelines in which different viperid clades diverged, helping to understand morphological diversification in a biogeographical context
[48]. However, these initial studies examined the evolutionary value of mitochondrial genes, revealing that genes such as cytochrome
b and NADH are the suitable predictors of phylogenetic relationships, largely due to the slower evolution of ribosomal unities
[49].
Initially, Parkinson et al.
[50] and Gutberlet Jr et al.[51] identified and suggested the existence of two monophyletic groups of pitvipers in America: A clade of North American species that groups
Agkistrodon,
Atropoides, Cerrophidion Crotalus,Metlapilcoatlus, Ophryacus and
Sistrurus, and a second clade that groups the other genera located in the Neotropical region. One analysis
[50] recovered Americas viperids as a monophyletic group but without a suitable identification of their sister group: a single invasion of North America with a subsequent divergence between the temperate and tropical north and at least three invasions of South America by the pitviper historical ancestors. Furthermore, one study
[50] reported that the species at the time considered as
Porthidium hyoprora corresponded to a lineage more closely related to the
Bothrops group (coinciding with the first description
[52], naming it
Bothrops hyoprora), resulting years later in the description of a new genus,
Bothrocophias that would include other species also included within the
Bothrops group, such as:
B. campbelli,
B. hyoprora,B. microphthalmus, and
B. myersi[8].
In Colombia, the significant elevation of the three Adean Cordilleras played a pivotal role in the cladogenesis of numerous pitviper species, such as the
Bothrops species group, currently distributed allopathically
[53]. The Andes Cordillera did not reach a maximum elevation of more than 40% until the Neogene era. This resulted in significant mountain formation only during the late Miocene and especially during the Pliocene, when the orogenic process accelerated quickly
[54]. Hence, it is posited that the initial division among the inter-Andean pitviper groups in northern South America is attributed to a dispersal event within the Caribbean of Mesoamerica that influenced the biogeography of the region.
Molecular studies of pitvipers have primarily focused on the Neotropical region, examining various hypotheses that could explain their astonishing diversity
[55–57]. Phylogenies have been used to explain the evolutionary trajectories that give origin to the types of plants and animals that currently live with us, helping to postulate theories that try to explain the diversity of organisms during geological time scales (e.g., during the Pliocene and Miocene periods) and after milestone geological events such as the Andean orogeny or the uplift of the Isthmus of Panama.
The uplift of the Andes in South America was, in fact, one of the most significant geological developments in the evolution of numerous vertebrate species
[58,59]. For instance, some authors utilize historical migration analysis in populations of the lancehead (B. asper) to examine its recent diversification events in South America
[60]. Their methodology includes the incorporation of multiple demographic processes into the inferences about patterns of divergence that are not feasible to infer or comprehend using methods based on phylogenetic trees
[61,62].
Indeed, the lancehead pitviper species have demonstrated that the impact of habitat diversity is correlated with phenotypic variability, a biological phenomenon inherent to the different
Bothrops species
[63,64]. The absence of any other potential adversaries at the onset of the expansion of distinct regions in northern South America may have contributed to the spread of the lanceheads across most of the geographical regions of Colombia. Nonetheless, it is imperative to emphasize that certain genera such as
Porthidium (P. nasutum and
P. lansbergii) and
Lachesis (L. acrochorda and
L. muta) exhibit convergence with
B. asper and
B. atrox in some ecoregions and localities. Nevertheless,
Porthidium species exhibit a widespread distribution in the southwestern zone of the Pacific region
[65], and in the north-Caribbean region
[66], being a species that are sympatric with
B. asper.
Nowadays, the bioinformatics methods have enabled the analysis of extensive datasets, such as genomic data, allowing the evaluation of intricate historical processes with high resolution
[67–69]. Furthermore, the genome has proven to be especially useful due to the accuracy and precision with which the different parameters are estimated
[70–72]. In summary, systematic deficiencies in certain pitviper studies have been addressed by utilizing integrative taxonomy and various lines of evidence for species delimitation, including morphological characteristics, genetic distance analysis, molecular phylogeny, and coalescent species delimitations that can encompass geographic and ecological isolation
[73–75].
2.1 Understanding how diversification events affect the diversity of pitviper species
Numerous hypotheses on Neotropical diversification have been proposed, including allopatric speciation and ecological displacement through mountain ranges
[58], rivers that functioned as prezygotic barriers, or habitat fragmentation caused by climate change during the Pleistocene
[76]. Continental drift of South America and the uplift of the Andes resulted in the evolutionary dynamics that underpin the foundation of diversity in numerous ecoregions of the Neotropic
[77–79]. These orogenic phenomena have raised questions about how many taxonomic groups were colonized and differentiated, and which ones would later inhabit the lowlands and slopes once the Andes reached their current elevations
[80]. In this way, pitviper diversity in South America is connected to biotic exchange between North and Central America throughout the Tertiary period, especially after the emergence of the Isthmus of Panama
[81].
Most speciation events in tropical regions are primarily influenced by the isolation induced by landscape alteration (e.g., vicariance) that is intrinsically linked to a broader range of biological and geological processes
[60]. Despite this, the current phylogeographic hypotheses proposed for South American pitviper species have not been evaluated using a consideration of the influence of demographic processes
[55,57,60,82]. In fact, examining the phylogeographic patterns of distinct species with overlapping or partially overlapping ranges can reveal common events that may have a similar impact on the evolutionary patterns of numerous taxa
[83]. However, inconsistencies among sympatric taxa may arise because of lineage sorting, variation in effective population size, extinction, dispersal, sympatric speciation, or non-response to vicariance events
[84–86].
In contrast, the explanation for parapatric speciation along the mountainous vertical axis could be attributed to specific adaptations associated with ecological gradients
[87]. Consequently, the capacity of certain species to expand into novel niches that emerged during a specific time must have been pivotal in the diversification of inter-Andean pitvipers belonging to the genera
Bothrops,Bothrocophias,Bothriechis,Lachesis, and
Porthidium. These events in northwestern South America during the early Pliocene suggest that the final uplift of the Andes could have played a significant role in the species diversification (=cladogenesis) of pitviper groups
[53].
Therefore, it has been posited that the rapid increase of viperids species from a common ancestor (with heat-sensitive pits) represents a significant evolutionary milestone, enabling the successful diversification of this family of snakes
[3]. From an ecological standpoint, these lineages inhabit diverse environments and habitats, ranging from tropical lowland forests in Central America and the Amazon to open areas in the Andes and in Patagonia
[2,55]. However, numerous species are still poorly known with respect to their taxonomy, natural history, and phylogenetic relationships
[2,28,57,88]. In fact, several groups of pitvipers show apparently endemic patterns, and because some taxa are considered species complexes, several species groups are in the process of redescription and resolution of their phylogenetic patterns (see Chapter 1).
One of the significant challenges faced by herpetology in Colombia is the limited number of molecular markers that have been examined in species of snakes. Furthermore, the limited knowledge regarding the rates of molecular evolution of various groups of snake genes precludes a comprehensive phylogeographic and historical examination of the pitvipers found in northern South America.
2.2 A family with distinct species complexes
In Colombia, as well as throughout the Andean ecosystems, the most significant environmental factors affecting the distribution of the seven known pitviper genera are elevation and orographic complexity. These factors are proposed as some of the main drivers of the phylogenetic relationships and venomic diversity in several pitviper species. For instance, a select group of Colombian pitviper species attain elevations ranging from 1,500 to 3,200 meters above sea level, with 1,500 meters being the upper limit for most pitviper species inhabiting the lowlands; 3,600 meters is the upper limit for high Andean species (such as
Bothrocophiastulitoi,
B. myrringae,
Bothriechis schlegelii, and certain populations of the
B. atrox complex)
[2,75].
Within the diverse biogeographic regions of northern South American, the proportion of venomous species can range from 9% to 14%
[89]. Therefore, the distributional zones of Colombian pitvipers have been primarily concentrated in areas inhabited by humans, in contrast to the notable presence of several snake populations in other forested areas
[90]. As a result, the dispersion of the Colombian pitvipers is associated with primary forested areas, secondary forested areas, areas intended for agriculture at different elevations, and increasingly with urban areas (usually in expansion). Consequently, and due to their wide range of geographical distribution in Colombia,
B. asper and
B. atrox are the most medically important venomous snake species
[64,91].
Historically, the genera
Bothrops and
Porthidium have been included in a substantial number of reviews and taxonomic approaches that have revealed a diversity previously unrecognized at the species level
[63,92–97]. Over the past two decades, some species belonging to the genus
Bothrops have been the subject of numerous phylogenetic studies and biogeographic approaches pertaining to their diversification on the southern side of the western hemisphere of the earth
[3,38,55,98,99]. These approaches have revealed a taxonomic complexity mainly within the
Bothrops, although currently work is also being conducted with the
Porthidium.
Research on the presence of cryptic diversity for several species of pitvipers found in Colombia reveals the presence of some species complexes identified through a series of common characteristics shared between several species and lineages (see Chapter 1)
[53,63,75,92–95]. This difficulty in distinguishing morphologically distinct species (cryptic species) that are genetically divergent has become one of the most discussed points in work published in molecular systematics and in the process of species delimitation.
One of the objectives of this section is to document some of the pioneering research on the systematics, venoms, and evolution of Colombian pitvipers. Based on published data over the past three decades, we believe it is necessary to increase phylogeographic studies in order to analyze the cryptic diversity of recognized taxa
[48,83,94,96]. This information could serve as an important tool for determining conservation efforts aimed at preserving viable populations of lineages that are yet to be discovered
[83].
Below we present a description of the main groups and species complexes of the medically important pitvipers in Colombia, and we review the principal works that have laid the foundations of our knowledge. We also discuss pitviper systematics. Finally, we compile other published information about the different groups to show the association between the studies carried out on Colombian pitviper venoms and their phylogenetic correlation. Thus, we intend to show not only the phylogenetic complexity of the two main groups (Bothrops and
Porthidium), but we also want to show the lineages and populations by species where there is no genetic or proteomic information.
The most medically important Colombian lancehead pitvipers: the Bothrops asper-atrox complex
The genus
Bothrops exhibits more genetic divergence in comparison to the other South American pitvipers. In fact, the South American
Bothrops exhibit paraphyletic characteristics in relation to the species found in Central America. The diversification of the genus has probably taken place in South America
[60]. This hypothesis suggests that the common ancestor of all
Bothrops was the first viperid to colonize South America sometime during the Miocene, about 10 million years ago
[63]. As a result, only one lineage of
Bothrops species (B. asper) spread throughout Central America, from Panama to Mexico. However, there are geographical distribution records for a second species of
Bothrops (B. punctatus) in the eastern region of Panama.
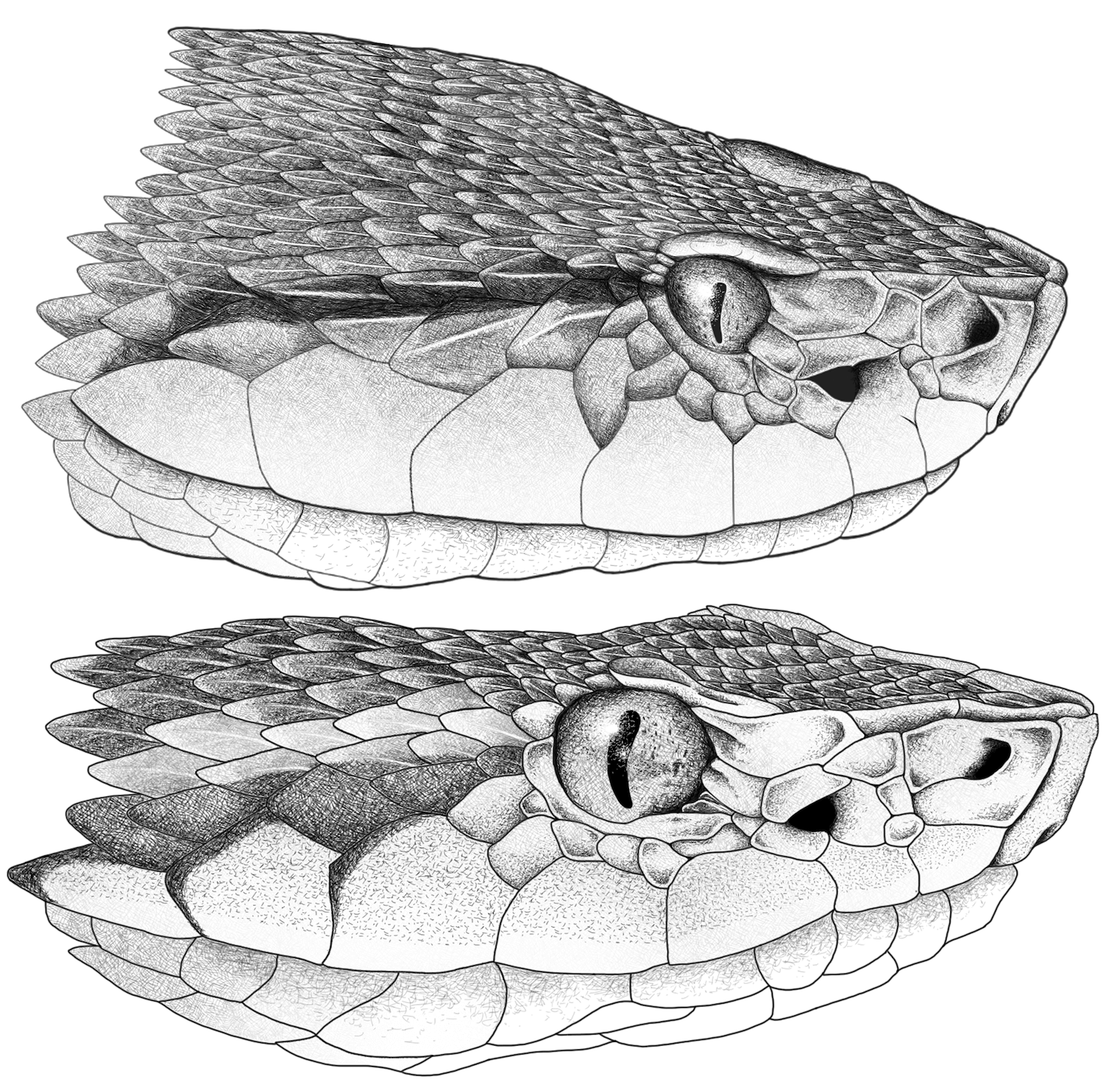
Figure 1. Head illustrations of
Bothropsasper-atrox complex. (Top): head in lateral view of
Bothrops asper from Melgar, Tolima, Colombia (INSZ 138). (Bottom): Head in lateral view of
Bothrops atrox from Puerto Carreño; Vichada (BOTATR00061). Illustrations by Oscar Ramirez.
Campbell and Lamar
[2] suggested that Colombian populations of
B. asper and
B. atrox species have allopatric distributions. In fact,
B. asper (Figure 1 Top) is distributed throughout the biogeographic regions of Choco and Magdalena Valley, with a focus on the Caribbean and inter-Andean basins
[64,91], whereas
B. atrox (Figure 1 Bottom) is found in the Orinoquia and Amazon regions. This distribution aligns with the dispersal hypothesis for the
B. atrox species, positing that a preexisting population of forests located on the northern side of the Amazon gradually colonized the regions bordering the Amazonian River
[97].
Nonetheless, there could be lineages of
B. asper and
B. atrox insympatric geographical regions of Colombia’s eastern and western Andes, like Antioquia, Cauca, and Choco. Several studies have demonstrated that both species show recent lineage differentiation in relation to habitat diversity as well as numerous unrecognized evolutionary lineages associated with elevational gradients
[53,60,92,97].
The taxonomic status of the
B. asper-atrox complex within Colombian populations has been little explored from a genetic perspective, and there is no clear delimitation between the lineages of these species to help recognize some of the previously described taxonomic hypotheses based on morphology. Particularly, the populations on the eastern slope of the Cordillera Oriental (B. asper-atrox complex) represent a challenging problem still unsolved: Some
B. atrox mountain populations have been historically reported as
B. isabelae[98] and
Bothrops colombiensis[99].
Bothrops isabelae was initially synonymized with
B. atrox in the 90s
[33,100], but later it was recognized as a species in some of the pioneering work for South American
Bothrops
[92]. Recent analysis
[101] shows
B. isabelae together with
B. atrox withan apparently low genetic distance. Even so, the authors recognize
B. isabelae as a valid taxon, although a few years later it was again considered a synonym of
B. atrox[55]. Campbell and Lamar
[2] consider
B. colombiensis as a species synonymous with the species complex formed by
B. asper and
B. atrox from Venezuelan populations.
Therefore, clarifying the phylogenetic connections between the Colombian
Bothrops species and the populations of Venezuelan species/lineages would enable us to comprehend the dispersal dynamics of numerous of these widely distributed taxa, providing an alternative to the utilization of the unified species concept widely applied in analyses of geographic lineages
[102]. The complexity of understanding the systematics of the
B. atrox group is challenging
[63,88,103]. Some studies suggest that the clade composed of
B. leucurus and
B. moojeni is the sister group of
B. atrox; other studies indicate that
B. atrox has closer evolutionary relationships with
B. asper and
B. isabelae[6,94,101,104]. Nonetheless, the
B. atrox group originating from Colombia and Brazil requires a detailed taxonomic review, as the taxonomic hypotheses based on morphology do not align with the described mitochondrial lineages. Furthermore, these mitochondrial lineages also do not correspond with the taxonomically recognized species
[103].
An example of this marked taxonomic complexity for
Bothrops in Colombia was the naming of new species derived from
B. asper[105]. That research provided a study that reevaluated the taxonomical boundaries within
B. asper populations in Colombia and proposed
B. ayerbei and
B. rhombeatus as new species within the
B. asper complex. Nonetheless, the unsuitable quantity and quality of characteristics employed in the species delimitation, the ambiguous descriptions of morphologic traits and diagnostic characters
[106], and the lack of a phylogenetic analysis, impedes a clear and objective distinction between the species proposed (see Chapter 1). In fact, in the initial description made by García
[107] no type specimen was assigned nor was the description of a new taxon made, turning
B. rhombeatus into a
nomen dubium (see Chapter 1).
Another crucial point mentioned by Ramírez-Chaves and Solari
[106] refers that Folleco-Fernández
[105] did not describe a neotype for
B. ayerbei and
B. rhombeatus nor were data on the geographical distributions provided for possible hybridization between the species nor was the relevance of distinguishing both taxa as valid species discussed. Although other authors have incorporated molecular markers to assess the phylogenetic relationships within the
B. asper-atrox species complex
[53]. Currently, the sequences from
B. ayerbei and
B. rhombeatus populations distributed in the southwest of Colombia (Cauca and Nariño departments), proposed by Folleco-Fernández
[105], are not available in official gene repositories, being lineages synonymous with
Bothrops asper and
Bothrocophias colombianus, respectively. So, further studies should include a larger or sufficient series of individuals collected in different habitats and analyzed with an integrative taxonomic approach in their sampling methodology
[60,97].
In order to highlight the absence of genetic and venomic information of some representative clades and lineages within the Colombian pitviper species, we present as a novelty a heat map that shows the phylogenetic relationships of the
B. asper-atrox complex and some proteome data published for some species in the last two decades. Thus,
Figure 2 shows the result of a molecular phylogeny constructed from fragments of two mitochondrial genes, and supported by some genera of pitvipers as out groups (Appendix). We have recovered a topology that includes terminals for different populations and lineages, as well as the recently described Colombian
Bothrocophias species
[75]. We used three terminals from three populations in Brazil, including two terminals reported as species but recently synonymized for the species
B. atrox. We used data from eight groups of protein/toxin families that have been frequently reported in venomic studies, and we performed the association of the percentages of these main components to present hypotheses related to the conformation of the clades and their geographical distribution (Figure 2).
We suggest that
B. asper from Colombia presents three large and well-supported phylogeographic lineages that are morphologically and ecologically diverse with genetic divergences between 2.5% and 4%. On the other hand,
B. atrox presents undefined population groups whose phylogenetic relationships still need to be well established (populations west of the Cordillera Oriental - Cundinamarca vs. populations east of the Cordillera Oriental- Meta, Vaupes and Caqueta). Furthermore, the geographical and taxonomic limits with its synonymous species/lineage
B. isabelae (also distributed in the eastern versant of the Cordillera Oriental in some parts of Venezuela) continue to be a taxonomic problem to be resolved. Here we use information from ongoing work to indicate that revision of the taxonomic status of the species/lineage
B. isabelae could be reported later.
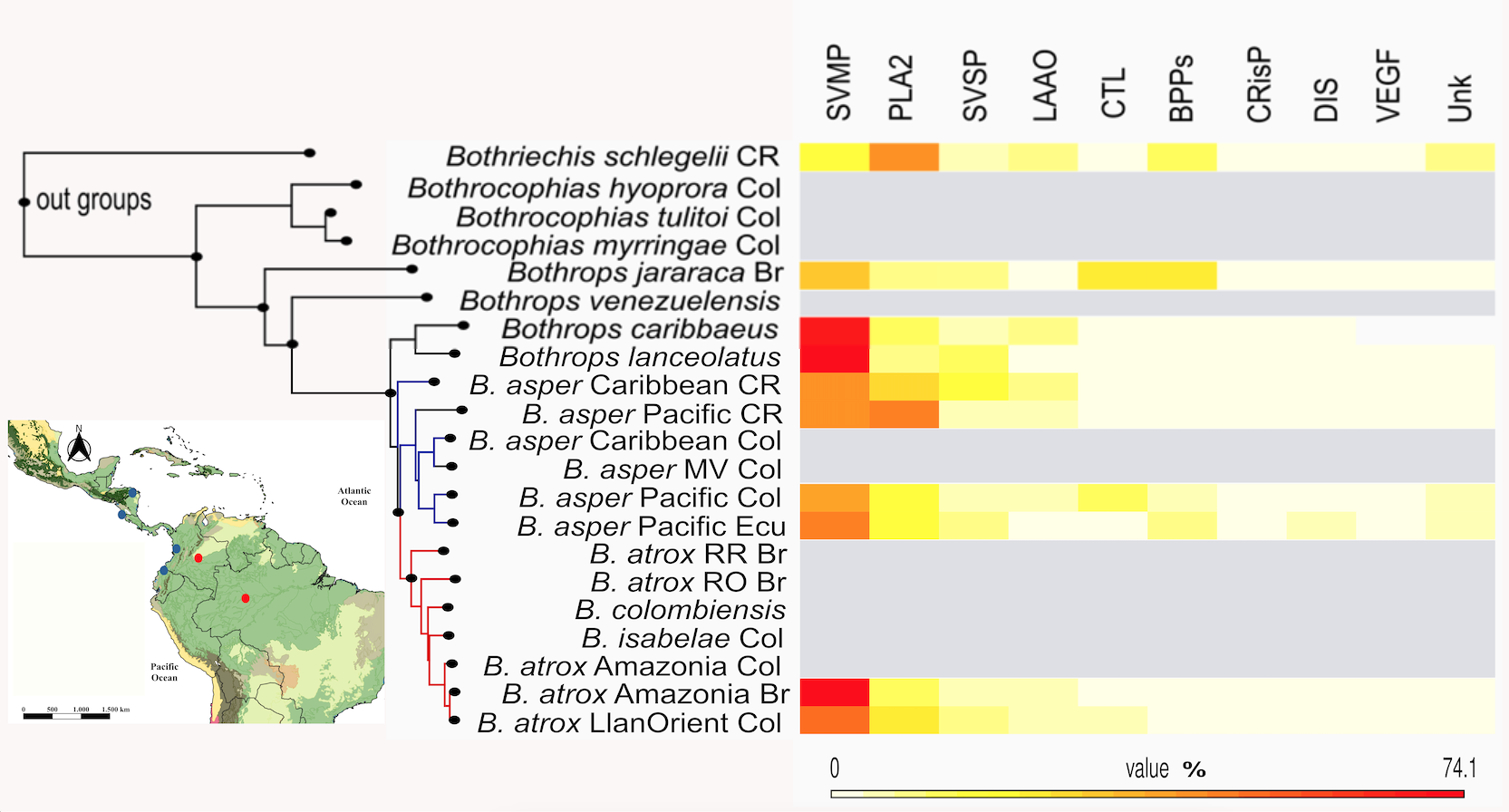
Figure 2. Molecular phylogeny of concatenated genes (MT-ND4 and MT-CYB sequences, produced by
[3,50,53,63,75,94,96] and Cubides-Cubillos et al. (unpublished data) using maximum likelihood (ML) and a proteome heatmap of the Colombian pitvipers (Bothropsasper-atrox complex). Phylogenetic topology retrieves the species and their most important lineages and those for which genetic and proteomic information is available (see bibliographic information in Section 3). Besides, the nodes that support branches with bs > 90% are shown. Abbreviations: lineages/populations (Caribbean and Pacific CR: Costa Rica; Caribbean and Pacific Col: Colombia; Pacific Ecu: Ecuador; MV Col: Magdalena Valley, Colombia; RR Br: Roraima region from Brazil; RO Br: Rondonia region from Brazil; Amazonia Br: Amazonian region from Brazil; Amazonia Col: Amazonian region from Colombia; LlanOrient Col: Orinoquia region from Colombia; and protein family names: SVMPs: metalloproteinase; PLA2s: phospholipase A2; SVSPs: serine proteinase; LAAO: L-amino acid oxidase; CTL: C-type lectin/lectin-like; BPP: bradykinin-potentiating peptide; CRISP: cysteine-rich secretory protein; Unk: nucleotidase; phosphodiesterase; hyaluronidase; nerve growth factor; peptides and/or non-protein compounds.
It is important to note that the sister genus of
Bothrops,
Bothrocophias, has only three species with genetic data and a single published proteome (see
section 3 in this chapter). Likewise, we were not able to calculate this relationship analysis for the proteomes of the
B. ayerbei and
B. rhombeatus lineages due to the absence of genetic data available. Finally, the absence of venomic data for two important lineages within
B. asper (populations from the Caribbean region and Middle Magdalena) is noted here.
With relation to the species
B. atrox and the various lineages found in previous phylogenetic analyses, only records of proteomic data were found for a population from Colombia (Llanos Orientales region, Meta department) and the northern region of Brazil (Amazonas State). It remains under consideration for subsequent analysis, whether the populations of
B. atrox distributed along the eastern versant of the Cordillera Oriental (encompassing Boyaca department), populations of the Colombian Amazon in the foothill region (Caqueta and Putumayo departments) of the Eastern Andes and those from the Amazon lowlands (departments of Guainia, Guaviare, Vaupes, and Amazonas), would require a review of their venoms, as well as other molecular data. Likewise, data from highland populations on the eastern versant of the Cordillera Oriental identified as the
B. isabelae lineage, could show certain types of venomic adaptations such as biochemical diversity, functional specialization, and evolutionary dynamics as gene duplications, diversification and rapid evolution of toxin genes, which represents a challenge in future research for mountain populations pitvipers genus
Bothrops.
The hemorrhagic potency of several types of Snake Venom Metalloproteinases (SVMP) is one of the important factors in the biological action of
Bothrops venoms. Its relative abundance in venoms is related to local and systemic hemorrhage
[43]. Our relationship analysis showed that the species and populations with the highest content of SVMPs were geographically distributed in both dry and humid tropical regions. Furthermore, venom composition exhibited high variability within closely related lineages, indicating a possible ecological response. Probably, the heterogeneous distribution and dispersal of species/lineages is a possible cause of the synergistic effects between several types of SVMPs. In fact, hemorrhage, which also contributes to myonecrosis
[108], is one of the most documented effects in snakebite accidents of all members of the
B. asper-atrox species group.
The PLA2, the second most abundant toxin in the
B. asper-atrox species venom is responsible for myonecrosis and is closely related to the digestive functions of the venom
[109-110] (see Chapter 5). This toxin usually represents about 20-40% of the proteome venom composition of the
B. asper-atrox species group but shows moderate variability. In fact, the variability in PLA2 percentage seems not to be related to a specific clade, region, or climate. However, due to large information gaps, this pattern could change in future studies or could be associated with ecological variables such as prey availability, habitat loss, or anthropogenic pressure.
Enzymes incompletely mimic the action of thrombin, and Snake Venom Serine Protease (SVSPs) exert a variety of actions on hemostasis and the kinin system or kinin-kallikrein system
[111,112]. They constitute the third most important group of toxins and, in fact, show minor variation within the species/lineages of the
B. asper-atrox group. Reports of snakebite accidents do not place much emphasis on variations in coagulopathies or inflammation, as these are typical symptoms of bothropic envenomation (see Chapter 9). Evolutionarily, this appears to be a characteristic intrinsically linked to constructive interactions with other components of the venom (see Chapter
5 and
9).
The Colombian hognose pitviper:
Porthidium species group
Hognose pitvipers inhabit Central and South America, and it is hypothesizedthat their dispersal occurred along the Pacific coast during the final phase of the emergence of the Panamanian isthmus (3.5 mya), even after its formation
[63]. This dispersion might have resulted from interspecific competition with other viper species such as
B. asper, a species that has been identified as another pitviper that colonized South America before the Panamanian Isthmus
[2,53,63]. The species of
Porthidium are short-bodied terrestrial snakes that do not exceed 100 cm in length and whose hypothetical evolutionary origin is related to Central America
[2,3,13,14,113].
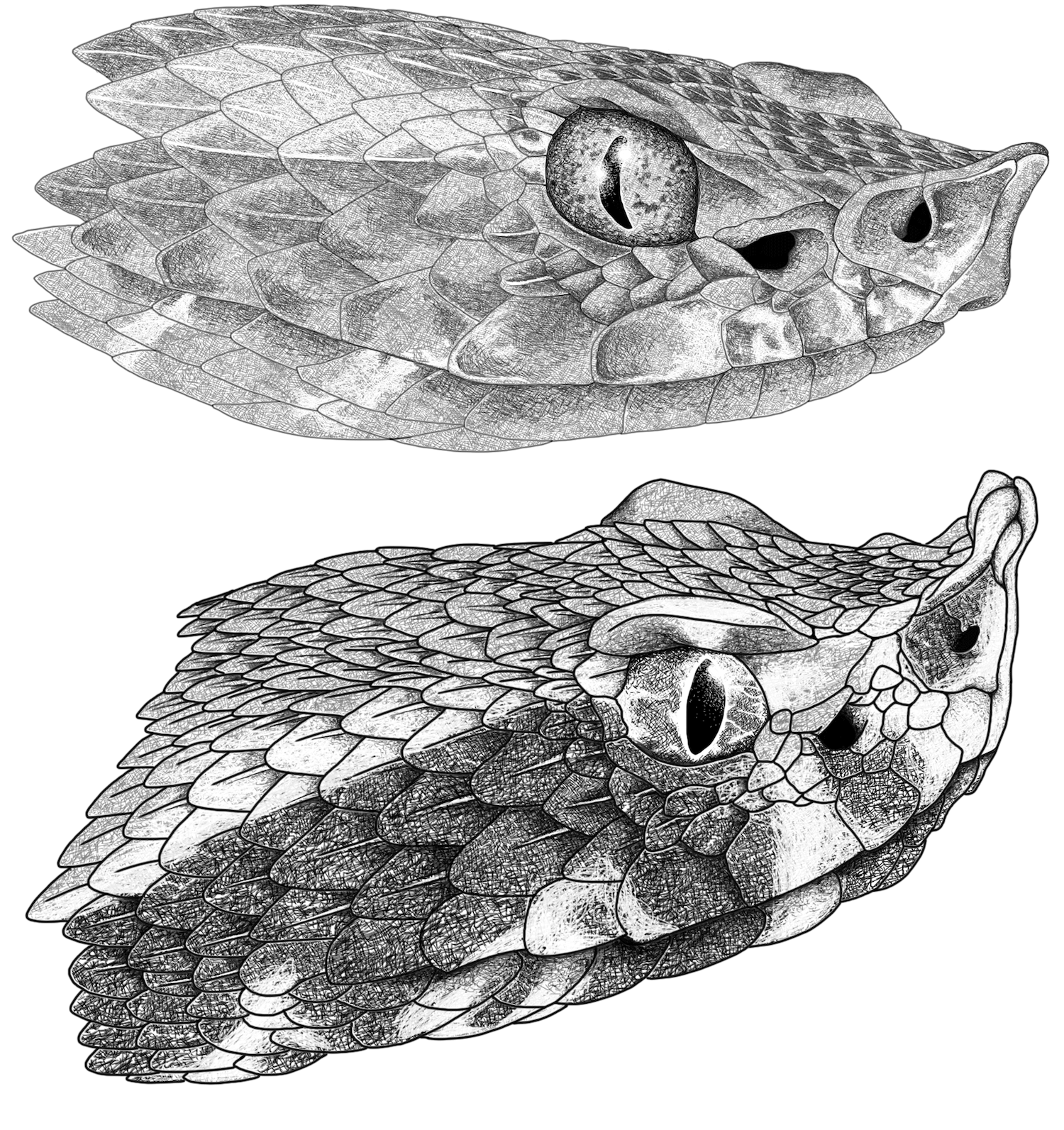
Figure 3. Head illustrations of
Porthidium lansbergii-nasutum species complex. (Top): Head in lateral view of
Porthidium lansbergii (INSV-SR-89) from Yondó, Antioquia, Colombia. (Bottom): Head in lateral view of
Porthidium nasutum (QCAZR 15395) from Tundaloma Lodge, Durango, Ecuador. Illustrations by Oscar Ramirez.
This genus shares cryptic dorsal color patterns and a clearly defined canthus rostralis (rostral upward scale; see Figure 3) that presumably denotes important adaptations for the hunting forms typical of ambush predators
[2]. In the 1990s, phylogenetic relationships between different species were initially reviewed using morphological characters and molecular data
[6,50,104].
Currently, various taxonomic studies have revealed the evolutionary and systematic relationships between the different species that support the taxonomic validity of nine species of the genus
Porthidium:
P. dunni,
P. hespere,
P. ophryomegas,
P. volnicacum, and
P. yucatanicum with a strong affinity towards arid zones; and
P. nasutum,
P. porrasi,
P. lansbergii,and
P. arcosae that inhabit lowland tropical rainforests
[95]. It is important to highlight that the description of the latest species for Central America,
P. porrasi[114], was based on an analysis of adjacent populations of
P. nasutum from Costa Rica. This implied the initiation of a discussion on phylogenetic diversity within the genus and marked the first strong revision suggested for northern South American populations
[95].
In Colombia,
P. nasutum[115] and
P. lansbergii[116] are closely related species, characterized by cryptic morphological traits such as the 'hog-nose snout' character.
Porthidium nasutum is distributed from Mexico through the Isthmian-Atlantic moist forests, transitioning into the Chocó-Darién ecoregion, as well as the biogeographic Chocoan ecoregion, including Ecuador
[2]. Meanwhile,
P. lansbergii, is distributed in the Isthmian-Pacific dry forests of Panama and the Colombian Caribbean, before reaching the tropical dry forest in northern Venezuela and the Chocó-Darién region, where it transitions into the Urabá humid forests and the evergreen and dry forest in the Magdalena Valley
[2,117].
Most of the previous scientific contributions for
P. nasutum and
P. lansbergii, performed phylogenetic approaches based on geographically restricted and small individual sampling
[117,118]. The taxonomic complexity observed in Panama and in the diverse Colombian populations warrants large-scale research. For example, Castoe et al.
[95] demonstrated that
P. nasutum is a paraphyletic clade that shows two genetically well-differentiated lineages in Costa Rica and Ecuador.
Similarly, in our phylogeny of
P. nasutum we recovered three principal lineages: Clade 1 and Clade 2 from Costa Rica and Ecuador, matching with previous results reported Castoe et al.
[95], while Clade 3 clustering specimens distributed from the northeast of the Colombian Andes, and those from the central-eastern region of Antioquia department. Yet, in our phylogenetic analysis, the Colombian population (central-eastern Antioquia) was recovered as a clade related with the different lineages of
P. lansbergii group. Previously, Cisneros-Heredia and Yánez-Muñoz
[119] suggested that the Ecuadorian population ought to be regarded as a novel species, distinct from
P. nasutum in Central America. However, our phylogeny does not include samples from the Chocoan populations of
P. nasutum (Colombian Pacific Coast) or from the upper Magdalena River basin (Huila department), which could limit the resolution of the phylogenetic relationships between the
P. lansbergii and
P. nasutum Colombian lineages, as well as the inferences derived from this.
Our molecular phylogeny highlights that
Porthidium lansbergii exhibits a wide range of diversification across diverse ecoregions encompassing elevation ranges from sea level to 1,200 meters. The Colombian lineages exhibit paraphyletic groups: Populations from the Darién-Panama are distinguished from those located in the Colombian Caribbean ecoregion, the Magdalena River basin, insular populations, and those to the east in Venezuela. Furthermore, certain genetic sequences associated with the two subspecies of
P. lansbergii (P. l. hutmanni and
P. l. rozei) were grouped in a polytomic clade, indicating a close association between the lineages of the Caribbean coast and Magdalena Valley with individuals analyzed from Venezuela, as also reported by De Arco-Rodriguez et al.
[118].
Figure 4. Molecular phylogeny of concatenated genes (MT-ND4 and MT-CYB sequences, produced by
[95,114] and Cubides-Cubillos et al.) using maximum likelihood (ML), and proteome heatmap of the Colombian pitvipers (Porthidium genus). Phylogenetic topology retrieves the species and their most important lineages, and those for which genetic and proteomic information is available (see bibliographic information in Section 3). Additionally, the nodes that support branches with BS > 90% are shown. Abbreviations: lineages/populations (CR: Costa Rica; Ecu: Ecuador; EasternAntCol: Eastern of Antioquia department, Colombia; Urabá Col: Urabá region from Colombia; MV Col: Magdalena Valey, Colombia; Caribbean Col: Caribbean region from Colombia; LlanOrient Col: Orinoquia region from Colombia; Darien Pan: Darien region from Panama; IslMarg Ven: Margarita Island from Venezuela); and protein family names: (SVMPs: metalloproteinase; PLA2s: phospholipase A2; SVSPs: serine proteinase; LAAO: L-amino acid oxidase; CTL: C-type lectin/lectin-like; BPP: bradykinin-potentiating peptide; CRISP: cysteine-rich secretory protein; Unk: nucleotidase; phosphodiesterase; hyaluronidase; nerve growth factor; peptides and/or non-protein compounds).
Only a few species of
Porthidium have published proteome composition. Despite of
Porthidium second genus of viperid with the greatest medical importance in Colombia (see Chapter 9), only one population
P. lansbergii has a proteome description
Nevertheless, the relationship analysis of molecular phylogeny and proteomic data of
Porthidium species shows that there are significant differences between the proteomes of sister species. However, due to the fragmentary state of knowledge about their venom compositions, it is not possible to propose an explanation for the apparent variability observed between the lineages, nor about the concordance between phylogeographic lineages and ecoregions, or ecological characteristics.
The main toxins that vary between species correspond to the three main toxin groups: SVMP, PLA2, and SVSP. Interestingly, these toxins vary in clades distributed in divergent climatic regions. However, missing data for several populations/lineages within the species
P. nasutum and
P. lansbergii could indicate different adaptive processes between populations of the same species. Therefore, we encourage both Colombian and foreign researchers to tackle biogeographic hypotheses that attempt to explain how the Andes promotes the random explosion of phenotypes (morphology, venom compositions, color patterns, etc.), as well as how selective pressure on venoms correlates or does not correlate with lineages and proteome variability
[188]. These hypotheses and speculations require future studies that significantly reduce the uncertainty surrounding the evolution of lineages within the
Porthidium genus and their relationship with the functional response of their venoms.
Giant Colombian pitvipers: The South American rattlesnakes and bushmasters
The South American rattlesnake
Crotalus durissus (sensu lato) is a remarkable species of
Crotalus; its venom is of significant medical concern due to its characteristic actions of neuromuscular paralysis, rhabdomyolysis, acute kidney injury, and coagulopathy
[120] (see Chapter
5 and
9). Recent research documents the diversity of subspecies for the species
C. durissus[121], reported as follow:
C. d. durissus
[11];
C. d. cascavella[9];
C. d. collilineatus[52];
C. d. cumanensis
[122];
C. d. marojoensis
[123];
C. d. maricelae
[124];
C. d. ruruima
[123]; C. d. terrificuss[37]; and
C .d. trigonicus
[125]. These authors include two additional species from northern South America,
C. unicolor
[126] with a geographic distribution restricted to Aruba and
C. vegrandis[127] with a geographic distribution restricted to Venezuela and previously regarded as a subspecies of
C. durissus (see Figure 5, Top).
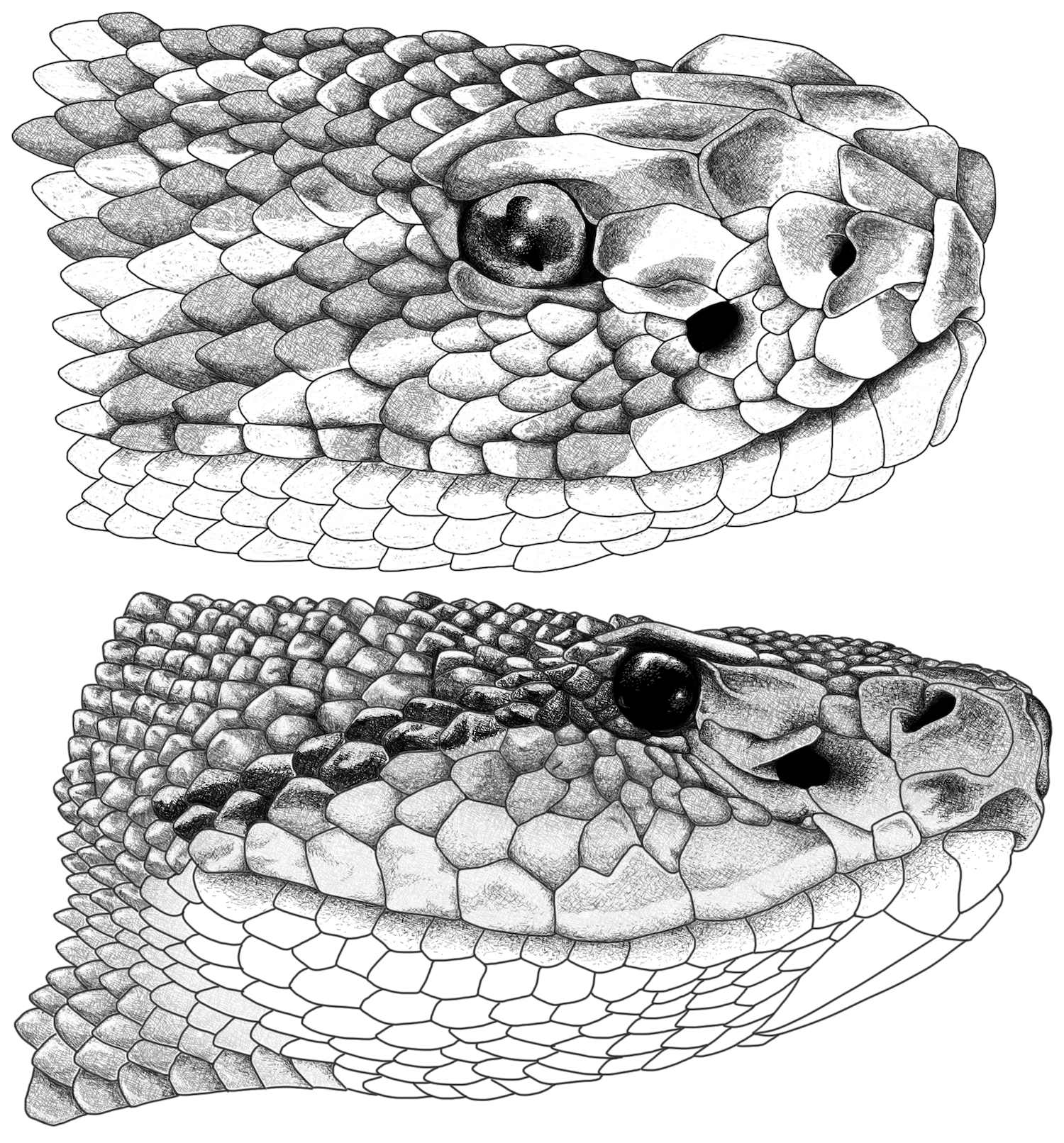
Figure 5. Head illustrations of giant Colombian pitvipers. (Top) Head in lateral view of
Crotalus durissus from the municipality of El Paso, Cesar Colombia (INSZ 115). (Bottom) Head in lateral view of
Lachesis muta from Mirití-Paraná, municipality of La Pedrera, Amazonas, Colombia (IAvH-R-8721). Illustrations by Oscar Ramirez.
Nonetheless, certain taxonomists have begun to recognize the subspecies as distinct species
[128,129]. In recent work, the
C. durissus group (including
C. vegrandis) has been phylogenetically recovered as a sister group to
C. simus (Central American rattlesnake)
[121]. However, it appears that the cryptic diversity in this group is the result of processes of isolation. Furthermore, the molecular phylogeny published for the
C. durissus group does not consider individuals from the trans-Andean and cis-Andean regions of Colombia.
In Colombia,
Crotalus durissus is distributed in the trans-Andean region of the country over the lowlands of the Caribbean region, extending into the Magdalena Valley, with disjunct populations in the middle and upper Magdalena River basin. In the cis-Andean region, it is found in the well-drained high plains of the Orinoquia
[2]. It is regarded as a species of terrestrial habits and is a prevalent resident of open areas that are relatively dry
[130]. his rattlesnake exhibits ecological flexibility, allowing it to be active both within forests and outside their edges, and it is commonly found in heavily human-altered open areas
[130,131]. Though it is one of the most venomous species in Colombia and one of the most illegally traded
[89], information on its natural history, registers of its occurrence, and evaluation of its state of conservation are still missing
[64] (see Chapter 1). The disjunct distribution pattern of its populations within the trans-Andean region, as well as between the cis-Andean and trans-Andean populations, suggests that it is very likely that
Crotalus durissus is composed of more than one evolutionary lineage that has diverged due to geographic and ecological isolation. Recently, significant intraspecific differences were found in the composition and biological activities of
C. durissus venom among populations distributed in the middle Magdalena River basin, the Llanos Orientales foothills, and the Colombian Caribbean coast (see Chapter 5). These results suggest that the hypothesis that
Crotalus durissus is a species complex with multiple evolutionary lineages that have diverged due to geographic and ecological isolation is plausible. However, future phylogenetic studies, including representative samples from all Colombian populations, as well as populations distributed in Brazilian ecosystems, are expected to help resolve these uncertainties.
The snakes of the genus
Lachesis[12], typically called “bushmaster”, are the largest pitviper snakes so far known, reaching more than 3.5 meters in length
[2,89]. The snakes of this genus are the only oviparous species among all American pitvipers
[132]. The species of the
Lachesis genus can be divided into Central American and South American lineages, with an estimated divergence between 6 and 18 million years ago. The divergence within the Central American species seems to have occurred between 4 and 11 million years ago, likely due to the uplift of the Talamanca Range. However, the differentiation within South American lineages occurred only about 800,000 to 300,000 years ago
[48,133]. Specifically, the emergence and structuring of the three Andean mountain ranges in Colombia has been proposed as the mechanism of isolation between
L. muta (restricted to the Amazon) and the Central American bushmaster species (L. melanocephala and
L. stenophrys), and
L. acrochorda, the only
Lachesis species inhabiting the trans-Andean region, distributed in the humid forests of the South American Pacific, the Darién region in Panama, and the middle Magdalena River basin.
Recent literature reviews for
Lachesis document the rarity of Colombian bushmasters, the number of individuals (specimens housed in biological museums;
see Chapter 1), a few facts about the nature of the genus, and the small number of snakebite accidents annually
[134]. In fact, it is surprising, given that
Lachesis is considered the largest venomous snake species in South America, reaching up to 350 cm in length
[89], and with a distribution encompassing four of the five Colombian ecoregions.
The systematics and taxonomy of the
Lachesis species are the best studied aspects of the genus. In fact, this research has explored hemipenial morphology, color pattern and scale counts. Despite of these Colombian populations of the two species distributed in the country are poorly represented in these studies or have not been included
[134].
It is important to highlight that in Colombia, the bushmaster species
L. acrochorda (see Figure 5, Bottom) and
L. muta have allopatric distributions: Populations of
L. acrochorda are distributed in the Chocó-Magdalena biogeographical province in western Colombia, and populations of
L. muta are distributed on the eastern slopes of the Cordillera Oriental and in the Amazon ecoregion
[135]. Colombian populations of bushmaster species have been poorly represented in phylogenetic assessments. For example, evolutionary relationships between the Colombian populations of
L. muta in the Amazon and the populations of
L. muta in the Atlantic Forest of Brazil have not been discussed.
Knowledge of the venomic and toxicological characteristics of Colombian bushmasters is still fragmentary. The protein composition of
L. acrochorda venom is known from populations from the Pacific Coast and middle Magdalena River basin in Colombia, allowing the appreciation that the conformation of
Lachesis venom is apparently very conservative in all species of the genus
[136,137]. However, in the future some studies of the molecular systematics and new venomic analyzes will be published among populations of
L. acrochorda from the southwest, northeast, and Pacific coast of Colombia (A.M. Franco-Vásquez personal communication 2024). For Amazonian
L. muta from Colombia few records are available; and therefore, no biological and natural history information has been documented
[138]. Thus, given medical importance of bushmaster populations from the Colombian Amazon, It is a priority to conduct venom proteomic characterization that allows to compare the venom compositions between populations from Colombia and Brazil.
Perspectives on poorly known species of Colombian
pitvipers
The last 20 years of herpetological research have allowed us to clarify several of the different biological processes for the main species of pitvipers in Colombia.
Bothrocophias and
Bothriechis represent one of the challenges for researchers working with genetic and molecular data. The recent description of two new species of
Bothrocophias reveals the importance of evaluating the cryptic populations of some species with a wide distribution (like
B. microphthalmus) and of a genetic approach to some groups of species present in several South American countries
[75].
Similarly, the proposal of new species
Bothriechis for Colombia opens a space for deeper research on the venoms of the only group of arboreal pitvipers in Colombia
[139]. Nevertheless, these efforts were hindered due to deep inconsistencies in species delimitation within Colombian populations, as well as the feeble evidence and unsupported new lineages proposed by Arteaga et al.
[139] (see Chapter 1). Recently, Reyes-Velasco
[222] re-evaluated the taxonomic proposal by Arteaga et al.
[139], demonstrating that these authors misinterpreted their results. The genetic differences they interpreted as species boundaries may instead reflect clinal variation, not independent lineages. The rest of the taxa proposed by Arteaga et al.
[139], including
B. khwargi,
B. klebbai,
B. rahimi,
B. rasikusumorum, and
B. torvus, do not hold up under more rigorous analysis and should be synonymized with
B. schlegelii. Therefore, we agree with Reyes-Velasco’s
[222] conclusions and recommend that future publications dealing with Colombian populations of
Bothriechis schlegelii refer to it as the
B. schlegelii species complex (see Chapter 1).
Published and available molecular data for different pitviper species inhabiting in Colombia were summarized in Table 1. However, despite several main phylogenies published on Neotropical pitvipers that provide our current understanding of the evolutionary paths of Colombian pitvipers, several taxa and populations of pit vipers in Colombian ecoregions are poorly represented or were not included in these phylogenies
[3,55,63,92,93,101,55]. Therefore, evolutionary paths of Colombian pitvipers still fragmentary.
Bothrops asper has been the only Colombian species extensively genetically tested from molecular supports using partial sequences of the cytochrome b and NDH4 genes
[53,60]. Also, parallel studies at the proteomic level could reveal the presence of geographical lineages for the main groups of species that are distributed North of South America, given that new research is being developed in the field of molecular systematics. We expect that the results of ongoing projects investigating the evolutionary trajectories of
B. atrox,
P. nasutum,
P. lansbergii, and
Crotalus durissus will arise in the coming years (Cubides-Cubillos et al.; and Vargas et al. in press), helping to unveil the complexity of the phylogenetic history of these Colombian pitvipers.
Table 1. Molecular marker data published for pitvipers from Colombia and other countries in the neotropical region.
(+) Data for Colombian species. (*) Data for species in other countries. (-) unpublished data. For the species
P. lansbergii and
P. nasutum, the molecular data correspond to research in progress developed by Cubides-Cubillos et al. as well as the data for
C.
durissus corresponding to a review developed by Mario Vargas et al.; the other sequences were developed by
[3,50,53,63,75,95,96,104] and
[114].
3. Biological activities and proteome of the Colombian pitviper venoms
The evolutionary selection of specific types of toxic components appears to have been somewhat limited. Therefore, it is posited that the venomic proteins belong to a limited number of protein families
[140]. Indeed, the restricted variety of proteins has undergone rapid evolution in its initial location, resulting in the production of numerous biological functions
[141]. Hence, as venoms are trophic adaptations that facilitate subduing prey and their effects are highly variable according to the type of prey and its availability, the diverse toxic secretions of snakes exhibit significant variation in their composition
[142].
Venom composition in snakes exhibits variation across all taxonomic levels (between families, genera, and species), yet understanding its mechanisms remains a subject of research
[143,144]. A plausible explanation for this variation may require the conduct of adaptation evolutionary studies, coevolution, and the establishment of a phylogenetic and ecological framework
[110,145,146,144]. The coarse description of the variability in venom composition has an impact on applied and biomedical sciences, influencing the efficacy of antivenom therapies and the treatment of snakebites, as well as the development of new drugs
[111,147] (see Chapter
6 and
10).
Several countries in South America exhibit a high prevalence of bothropic snakebites associated with serious and life-threatening envenoming (see Chapter 9). This has led to the stigmatization of the remaining pitviper species, as well as similar non-venomous snakes, resulting in their deliberate killing by diverse human communities across their distribution ranges. Nonetheless, apprehension regarding pitviper species belonging to the genus
Bothrops is not unfounded. Indeed, due to their capacity for inhabiting diverse ecological zones,
Bothrops species are among the most dangerous venomous snakes in tropical regions of Central and South America. Particularly in Colombia, bothropic envenoming accounts for 62% of annual cases
[111,148] (see Chapter 9).
Current research, focused on the study of venoms employing a multidisciplinary approach, using “omics” technologies such as genomics, transcriptomics, proteomics, and, more recently, metabolomics is used to investigate low molecular weight components
[149] of venoms. These research fields have had a significant impact on current biomedical science
[150]. Hence, by incorporating advances in high-resolution mass spectrometry (MS)
[151] and de novo and database-dependent annotation methods that need minimal amounts of venom
[152], enables identification of toxin families, individual toxins, unique isoforms, splice variants, amines, amino acids, and alkaloids.
More than a hundred Neotropical pitvipers are recognized worldwide
[4]; and given their extensive distribution, they constitute a medically important event with a significant impact on health, since they cause more than 2.7 million snakebites per year
[108]. However, snake venoms, at the same time, are a source of raw materials for the acquisition of new molecular compounds
[153], which facilitate various biotechnological advancements in molecular biology and biomedicine (see Chapter 10).
Chippaux et al.
[154] paved the way for the study of snake venom, showing that these multifaceted secretions can differ at different levels and are affected by various intrinsic and extreme factors
[144]. Currently, studies in venomic biology serve as models for studies in evolutionary genetics
[155,156]. In the following sections, we will provide a summary of the knowledge about venomics of Colombian pitvipers. We will also discuss the gaps and research perspectives achievable in the near future.
3.1 The complexity of venom: more than a hunting weapon
Pitviper venom contains a complex mixture of proteins and enzymes with different biological activities
[157]. Their primary function is to immobilize, pre-digest, and kill the prey (see Chapter 5). This enabled the transition from a mechanical subduction mechanism (body pinning and constriction) to a chemical subduction mechanism (the use of venom)
[158]. Thus, the venom system became a main feature of advanced snakes
[159], and it is an important trait for understanding the evolution and ecology of various snake species
[141].
These complex mixtures of proteins are produced by specialized buccal glands that are homologous to the Duvernoy and salivary glands and are also present in other members of the Colubroidea superfamily
[157,160]. These toxins originated from duplication events of genes encoding proteins with normal physiological functions (whose resulting proteins are recruited by the venom gland) and that show a selective expression on the structure
[141,161].
Over time, venom gland-encoded proteins were subjected to selection pressures that resulted in their structural and functional modification and even suppression of their expression in the gland
[161]. However, information about changes in the structural components and expression, as well as gene duplication related to the origin of the actual functions of various proteins and toxins in general, is still not well understood. Thus, understanding the order of such events that explain the origin of any protein is a challenge yet to be resolved
[162].
Snake venoms present great intraspecific and interspecific variability, largely because these are ecological traits that evolve dynamically. There are several factors that influence their synthesis such as seasonality, geographic distribution, ecological variation between populations, and the variety in the diet that intrinsically diverges with the age/sex of the individual
[154,163,164]. Since the properties of these venoms are useful when defending against predators, human beings have become part of this interaction.
For example, venoms from three clades of spitting cobras have the ability to activate mammalian sensory neurons, inducing pain
[165]. In fact, the divergence of the spitting cobra clades in Africa (6.7 Mya) could be associated with the evolution of bipedalism and the development of larger brains in hominids
[165], indicating that “spit venom” could be a mechanism that could have evolved in response to interactions with ancient hominins. In this sense, snakes will need to produce copious amounts of venom, which would implicitly make it difficult to infer whether the venom has a high-energy cost when compared to other ecological traits. In fact, it is difficult to determine experimentally this earlier premise due to the difficulty of experiments that would require specific prey management, taxon-specific toxicity testing, and, furthermore, an autonomous way in which snakes can modulate venom production and injection
[166].
In fact, today those circumstantial or provoked events in which a snake bites a human has been continued, technically called “snakebite accidents”
[167]. Hence, this incident leads to an intoxication resulting from venom inoculation from snakebite, resulting in a clinical case that is regarded by the World Health Organization (WHO) as one of the numerous neglected tropical diseases (NTDs;
see Chapter 9). Unfortunately, these snakebite cases occur more often in developing countries, where almost 95% of the total number of snakebite cases worldwide are reported
[168]. Moreover, since these accidents are frequent in low-income rural communities with limited access to healthcare services, addressing these envenomations is challenging due to the limited availability of antivenoms and the high costs of transporting the patients
[168,169] (see Chapter 6).
In perspective, venoms are a rapidly evolving trait; it has been inferred that environmental factors can generate adaptive pressures that modulate their functional variation across species. As an adaptive characteristic used both for feeding and for deterring or defending against potential predators, developing effective antivenoms to neutralize their envenomations is currently a significant challenge, since these therapies must account for the wide intraspecific divergence in venom composition. In fact, recent research that reports on divergence of crotamine content in populations of the genus
Crotalusdurissus should be a warning about the need to develop an antidote capable of neutralizing this toxin
[170,171].
One of the main “targets” of the venom of some snakes is the nervous system of the potential prey, even of one possible predator
[172]. The presence of neurotoxins in these venoms is attributed to their capacity to disrupt the normal functioning of the central nervous and/or peripheral nervous system
[173,174]. The high specificity and selectivity of neurotoxins found in rattlesnake venoms have been widely used to study the structure-function of receptors and other elements involved in neuromuscular transmission, neurological mutations, and the causes of various neurological conditions
[175] (e.g., Parkinson and Alzheimer,
see Chapter 10). However, today, little research is available on the effect of toxins on the peripheral nervous system and some specific organs
[172]. Researchers will continue to gain understanding about the targeted administration of drugs for the treatment of various nervous conditions, tumors, and neurodegenerative diseases in the future. In the interim, the persistent pursuit of research aimed at identifying novel ophidian peptides and comprehending their structure-function correlation with cellular receptors, will ultimately lead to the discovery of distinct molecules that have the potential to be utilized in diverse biomedical therapies (see Chapter 10).
In Colombia, the colonization of new niches in different mountainous, dry lowland, or humid tropical ecosystems must have played an important role in the phenotypic diversity of pitviper venoms that could be shaped by local adaptation to different habitats within ecoregions (e.g., the intraspecific differences in the venom of
Crotalusdurissus[171], see below). Nevertheless, further research efforts are still needed to understand the relationships between the composition of venoms, their ecological context, and their link to habitat transformations caused by human activities. For this, it is necessary to develop a more robust conceptual framework that allows for the proposal of hypotheses to explain the differentiation or specificity of venoms in an ecological and evolutionary context.
Understanding how these venoms have evolved in relation to interspecific interactions with humans would help to investigate the anthropogenic effect on natural populations of pitvipers, as well as to resolve some questions related to the effectiveness of antivenoms. Our goal in this section was to explain that snake venoms, before being a biological component that triggers a complex clinical picture (see Chapter 9), are a lethal hunting weapon modified and selected through the course of evolution of many species that grants the individual one of the most sophisticated natural resources for food and defense.
3.2. Studies in proteomics in Colombia
Despite the great diversity of pitviper snakes in Colombia, only nine species (40%) have been the object of proteomics studies of at least one population across their distribution range in the country (Table 2). The proteins present in these venoms are categorized into seven to twelve protein families. Generally, PLA2s are the most abundant components identified, followed by SVMPs, SVSPs, and LAAOs. However, there are some exceptions. For instance, the venom of
C. durissus exhibits a significant concentration of crotoxin (64.71%), which is a characteristic of South American rattlesnakes
[170,176], but between Colombian populations there are intraspecific differences in which crotamine is only present in the Caribbean ecoregion
[171].
Table 2. Percentage of toxins in the venom composition of the pitviper species distributed in Colombia with proteome characterization. Each toxin is expressed in its relative proportion (%).
| Toxins |
B. atrox |
B. asper |
B. ayerbei** |
B. rhombeatus** |
B. punctatus |
P. lansbergii |
B. myersi |
L. acrochorda |
| SVMPs |
48.5 |
33.17 |
53.7 |
39.7 |
41.4 |
35.5 |
21.50 |
23.2 |
| PLA2s |
24.1 |
31.29 |
0.7 |
23.0 |
9.3 |
16.2 |
54.04 |
2.3 |
| SVSPs |
10.9 |
3.89 |
9.3 |
4.9 |
5.4 |
4.5 |
3.43 |
35.1 |
| LAAO |
4.7 |
3 |
3.3 |
2.1 |
3.1 |
3.6 |
1.10 |
9.6 |
| Hya |
* |
* |
* |
* |
* |
* |
0.01 |
* |
| Crotoxin |
* |
* |
* |
* |
* |
* |
* |
* |
| CRISP |
2.6 |
1.45 |
1.1 |
0.7 |
1.2 |
1.4 |
* |
0.9 |
| Dis |
1.7 |
3.27 |
2.3 |
4 |
3.8 |
12.9 |
0.62 |
* |
| PLB |
* |
* |
* |
* |
* |
0.7 |
0.37 |
* |
| PDE |
* |
* |
0.7 |
0.2 |
* |
0.3 |
0.08 |
* |
| CTL |
7.1 |
8.54 |
10.1 |
3.1 |
16.7 |
6.7 |
0.56 |
6.9 |
| VEGF |
* |
* |
* |
* |
1.7 |
2.2 |
* |
0.6 |
| NGF |
* |
* |
0.1 |
* |
* |
* |
0.07 |
* |
| PNP |
* |
* |
* |
* |
* |
* |
* |
* |
| BPP |
0.3 |
* |
8,3 |
7 |
* |
* |
* |
21.5 |
| PEP |
* |
* |
8.7 |
* |
10.7 |
* |
9.07 |
* |
| Nuc |
* |
* |
* |
* |
* |
0.4 |
0.02 |
* |
| SVMPI |
* |
* |
* |
10 |
* |
* |
* |
* |
| Crotamine |
* |
* |
* |
* |
* |
* |
* |
* |
Abbreviations for protein family names: PLA2s: phospholipase A2; SVMPs: metalloproteinase; LAAO: L-amino acid oxidase; CTL: C-type lectin/lectin-like; CRISP: cysteine-rich secretory protein; SVSPs: serine proteinase; Nuc: nucleotidase; PDE: phosphodiesterase; Hya: hyaluronidase; NGF: nerve growth factor; PLB: phospholipase B; PNP: peptides and/or non-protein compounds; BPP: bradykinin-potentiating peptide. * Toxins no detected in the proteome. ** Dubious species of the
Bothrops asper species complex (see Chapter 1)
[118,136,171,181,184,187,192,195].
Until now, the venoms of
Bothrops species had higher proportions of SVMPs than PLA2. This same pattern was observed in the venoms of
P. lansbergii and
L. acrochorda. In contrast, the venom of
Bothrocophiasmyersi has a superior percentage of PLA2s. In addition, the venom of this species is the most complex, with twelve families of proteins. Special characteristics of
L. acrochorda venom are their elevated amounts of SVSPs and BBPs (35.1% and 25.5%), even more than PLA2s, which is a unique pattern. While the venom of the
L. muta (Amazon bushmaster) has the highest quantity of LAAOs with 9.6%.
Interestingly, the venom of
B. punctatus exhibits a higher concentration of C-type lectins (16.7%); however, these toxins were identified in all venoms through proteomic studies, ranging from 0,56% to 10,1% in the remaining species with proteomes available. It should be noted that the venom of
P. lansbergii possesses the highest concentrations of desintegrins, whereas these toxins were not detected in the venom of
L. acrochorda. One noteworthy discovery was the presence of hyaluronidases, the spreading factor of venoms (as discussed in
Chapter 5) that were only detected in the venom of
B. myersi, but in a minimal amount (0.01%). This result may be attributed to the fact that in certain instances not all proteins were classified. On average eight groups of toxins were described in most of the research, where percentages were usually informed for the following toxins: SVMPs, PLA2s, SVSPs, LAAO, CRISP, Dis, CTL and BPP. Crotoxin and crotamine are exclusive components for the
Crotalus genus.
The venom of
Crotalus durissus requires special attention because it is the only one with neurotoxic activity and low molecular weight myotoxins among Colombian viperids. The venom of Colombian rattlesnakes is composed of a considerable proportion of crotoxin (64.71%), the toxin responsible for inducing diaphragm-flaccid paralysis. However, another exceptional toxin present in this venom is crotamine, a small basic polypeptide with myotoxic and cell-penetrating activities
[177,178]. Interestingly, the expression of crotamine can be different in venom from different geographical regions, showing an intraspecific variation in the composition of the
C. durissus venom
[171,179].
3.3 The biological activities of Colombian pitvipers
All venom activities evaluated for species from Colombia are correlated with the signs and symptoms observed in snakebite cases reported (see Chapter 9). Venoms from
Bothrops species (B. atrox,
B. asper,
B. ayerbei,
B. rhombeatus and
B. punctatus;see Figure 6) have different lethal activity values (LD50 µg/mice), such as 81.4 (80.2–83.6)
[42], 63 (50–81 ranges for several ecoregions from Colombia)
[180], 50.1 (37.5–58.3)
[181], 54.9 (36.0–83.8)
[181], and 47 (36–61)
[180]. Hence, the most lethal venoms among the
Bothrops species tested so far are
B. asper and
B. punctatus. It is important to mention that not all Colombian
Bothrops species have been evaluated. However, the venom of
C. durissus is the most lethal venom within pitvipers in Colombia, with a lethal dose average of 1 µg/mice (0.02–2.5)
[171,180]. This lethal activity value is considered a consequence of the venom's neurotoxicity, a widely described functional feature that is biologically absent in the venoms of other pitviper species
[182,183].
Saldarriaga et al.
[42] carried out one of the pioneering studies on the ontogeny of
Bothrops venoms that allowed us to understand how the species in its first stages of life develops venoms with fractions endowed with high hemorrhagic activity, while in the adult stage mostly fractions are found with indirect hemolytic activity. Furthermore, when
B. asper venoms from Colombia and Costa Rica are compared, there are specific divergences associated with variations in lethal, hemorrhagic, edematogenic, myotoxic and indirect hemolytic activities
[42].
.png)
Figure 6. Biological activities of venoms of Colombian pitviper species (A) Lethal dose.(B) Coagulant activity. (C) Defibrinogenating activity. (D) Edematic activity. (E) Hemorrhagic activity. Acronyms: LD= Lethal dose of venom. MCD= M inimum coagulant activity dose of venom. MDD= Minimum defibrinogenating activity dose of venom. MED= Minimum edematic activity dose of venom. MHD= Minimum hemorrhagic activity dose of venom
In this study, we compared the biological activities of the principal biological important pitviper species from Colombia. We found that within the
B. asper lineages (B. asper,
B. ayerbei, and
B. rhombeatus[184]) the venom of main lineage of
B. asper (distributed throughout the Caribbean region and the middle Magdalena Valley) is significantly more lethal than
B. ayerbei, and
B. rhombeatus lineages.
Bothrops asper venom turned out to be more hemorrhagic than venom of the
B. rhombeatus lineage, but the minimum hemorrhagic dose of
B. ayerbeilineage was significantly lower, showing its greater capability to cause hemorrhage.
On the other hand, the venom of
B. rhombeatus lineage showed a greater coagulating capacity while
B. ayerbei lineage exhibited the lowest coagulating capacity. Moreover, despite the geographic isolation between the populations of
B. asper from Gorgona Island (Pacific Ocean) and the continental populations from the Valle del Cauca, the biological activities of their venoms showed very similar activities, with the exception of coagulating activity, which is much lower in the populations from Gorgona Island. However, the biological activities of the venom of these two populations are significantly different when compared to the venoms from the Caribbean cost and Magdalena River Basin
[185]. It has been inferred that the divergence of populations within some lineages of
B. asper is strongly influenced by the orography of the Andes mountain range, and the dynamically changing nature of their ecosystems that could drive specificity in different ecological niches
[184]. In fact, a proteomic comparison of
B. asper venoms based on the genetic diversity documented for lineages of the
Bothrops asper-atrox complex found intraspecific variation in four of the main components of the venom
[53,60]: SVMP, SVSP, PLA2 and C-type lectin-like proteins (CTL)
[184].
There is controversy regarding the biological activities of the venom of various populations of the
Bothriechis schlegelii species complex. As we explained earlier, this species exhibits clinal variation among its populations, which is possibly related to the variability in the biological activities of their venoms (Figure 6A, C-E). For example, the Colombian populations from the central-eastern Andes (Antioquia department)
[180,186] and those from the southwestern region of Colombia (Valle del Cauca department)
[187] do not exhibit hemorrhagic activity. However, the venom of these populations shows coagulating activity and a high capacity for edema formation, similar to what is observed in the populations from Costa Rica (Bothriechis nigroadspersus)
[188]. Given the medical importance of this species and its wide distribution along the agricultural production zones of the Colombian Andes, it is a priority to conduct rigorous studies exploring the determinants of the variability in the biological activities of the
Bothriechis schlegelii species complex. This will enable physicians treating envenomations to generate more accurate diagnoses, establish timely and appropriate therapy, and anticipate possible clinical complications.
Hemorrhagic activity of snake venoms is attributed to the activity of SVMPs
[189]. All pitviper venoms tested up till now have hemorrhagic activity; however, their potency is variable. Generally,
Bothrops spp.,
Porthidium spp.,
L.acrochorda and
Bothrocophias spp.are more hemorrhagic than the
B. schlegelii species complex, whereas
C. durissus rarely induces local hemorrhage
[190–193]. Nevertheless,
Bothrocophias campbelli venom induces mild hemorrhage and coagulation disorders
[194].
Both
Porthidiumlansbergii and
P. nasutum show medium cytotoxic activity and low lethality when assessed on mice, but both species show high hemorrhagic activity
[195]. In addition, pitviper venoms from Colombian populations induce hemostatic disorders that are related to their pro-coagulant activity shown in vitro, their fibrinogenolytic displayed in vivo, and the alteration of coagulation times. These venoms are characterized by a massive consumption of fibrinogen that can only be recovered by antivenom therapy
[90,193,196].
Table 3. Biological activities tested for Colombian Viperidae
* Dubious species of the
Bothrops asper species complex (see Chapter 1).
[42,118,136,171,180-190,194]
3.4 Outlook for antivenom research
Antivenom therapy, (based on serum therapy;
see Chapter 6), was developed in the 19th century; and although it has had huge improvements throughout its existence, this essentially uses immunoglobulins as therapy to this day
[197]. In fact, since their production, antivenoms have been used successfully for the treatment of snakebite envenomation in various regions of the world
[108]. In context, the antivenom “works” from purified immunoglobulins, which act initially by activating the immune response to the most pathogenic venom proteins, and then, trying to reduce and stop the severity of the envenomation. However, in some cases this therapy can cause adverse reactions (e.g., anaphylactic shock)
[111].
The administration of antivenoms is done intravenously. Once in the bloodstream, the antibodies or antibody fragments bind to the venom toxins, preventing them from damaging tissues or interfere with critical bodily functions. These immunoglobulins are obtained and purified from the plasma of large mammals (usually horses), which have been previously immunized with sub-lethal doses of snake venom
[198,199] (see Chapter 6). Antivenoms can be either monovalent or polyvalent, depending on whether the horses are immunized with venom from a single species or with a mixture of venoms from multiple species, respectively
[200].
The incidence and severity of adverse reactions associated with the administration of antivenom have been reduced by the purification of immunoglobulins
[201], thus demonstrating the therapeutic efficacy of antivenom to control the systemic manifestations of snake envenoming
[202]. However, the venom heterogeneity between species could explain the differences in the clinical symptoms recorded from different geographic regions during snakebite cases
[154]. This phenomenon reflects a series of local adaptations that confer a series of biological advantages on a certain population of snakes. Currently, snake envenoming represents a huge challenge for the design of new approaches to immunotherapy
[111].
The description of venoms from various species and the knowledge of implicit inter- and intraspecific variability raises a scenario of new and necessary research approaches for the selection of phylogeographic regions of interest to help capture and describe venom variability. For example, neutralization measurements of the
C. durissus venom using National Institute of Health of Colombia (INS) antivenom to calculate the median effective dose (ED50) have shown that more antivenom is required to neutralize venom from Caribbean populations (2.3 mg venom/mL) than those from middle Magdalena basin (1.4 mg venom/mL), or the Orinoquia ecoregion (0.6 mg venom/mL)
[171]. Therefore, there is a need to use venoms combined as a substrate to produce antivenom and, thus, prioritize the investigation of the snake venom using a combination of proteomic tools, functional toxicological and biochemical
[199]. In fact, Juan Calvete et al.
[111] presents a useful protocol to investigate antigen-antibody immunoreactivity, opening the door to the possible therapeutic utility of antivenoms against homologous and heterologous venoms in pitviper species.
Thus, based on the immuno-reactivity of the various toxins against antivenoms, these toxic components could be grouped into three types of toxins: completely immunodepletable, partially immunodepleted, and non-immunodepleted. Thus, immunization protocols must be improved by using immunogen mixtures
[111]. This approach has recently established the basis for the development of antivenoms on an immunologically sound foundation and has proposed the use of immunochemical analysis based on proteomics for the identification of the most medically relevant venoms from snakes present in a specific region
[111].
A first example of this type of new therapy, which has impacted and driven research in the field of antivenoms, is the development of an antivenom designed for the treatment of snakebites caused by
Bothrops lanceolatus (endemic to the island of Martinique in the Lesser Antilles). This Sanofi-Pasteur Bothrofav®, has demonstrated good preclinical efficacy in neutralization, timely preventing the development of the most severe systemic events
[203,204].
The limited availability of antivenoms in several Central and South American countries, as a result of various anthropogenic processes (social and environmental;
see Chapter 6), has created the need and challenge to improve the neutralizing capacity of the available antivenoms. As an alternative, the use of small molecular inhibitors has been proposed to counteract the effect of the main snake venom proteins that cause clinical symptoms. Research in this field has recently gained relevance
[144]. The effect of these inhibitors has been studied both in vitro and in vivo, yielding promising results for the inhibition of PLA2, SVMP, and SVSP
[205,206].
Researchers from the Clodomiro Picado Institute, led by Dr. José María Gutiérrez, conducted detailed proteomic studies of the venoms of
Bothrops caribbaeus and
B. lanceolatus (species included within the
B. atrox complex), finding immunoreactivity to the polyvalent antivenom produced in Costa Rica to neutralize the venom of Central American rattlesnakes. This antivenom was able to immunodegrade 80% of the proteins and neutralize the lethal activities of the venoms in these two species
[205]. Other studies highlight the importance of comparing proteomes between species with a similar natural history but with taxonomic uncertainties or undefined lineages within the
Bothrops asper-atrox complex
[111,207,208]. The findings of the Costa Rican researchers represent a significant advancement in understanding the cross-reactivity of antivenoms that counteract the symptoms of envenomation caused by American pit vipers.
In Colombia, experiments have been conducted to explore complementary options to antivenom treatment. Recently, recombinant proteins have been produced to improve the immunogenicity of proteins such as PLA2 and three-finger toxins, representing a novel advancement for the future development of antivenoms that neutralize the toxins of snakes from the
Micrurus genus
[209]. Additionally, experiments using purified molecules from plant extracts to inhibit certain biological activities of snake venoms
[192,210,211], as well as synthetic inhibitors that have shown inhibitory activity against PLA2 and SVMP
[212] have all been carried out in preclinical trials.
The gradual incorporation of chromatographic and mass spectrometry techniques to complement the current platforms recommended by the WHO in the evaluation of antivenoms are ideal for establishing a geographical range of their clinical validity
[213]. Besides, when combined with
in vivo neutralization assays, antivenoms represent a good complement for evaluating the therapeutic efficacy of antivenoms against the same venoms used in its production
[214].
Polyvalent antivenoms such as those from ICP (Costa Rica), INS (Colombia), PROBIOL (Colombia), UCV (Venezuela), BIOL (Argentina), and INS-PERÚ (Peru) can recognize the main toxin families present in
Bothrops asper venoms (SVMP, PLA2s, CRISP, SVSP, CTL); among these, the INS antivenom is the most effective and safest therapy
[181]. In comparative studies of the antivenoms available in Colombia, the INS antivenom showed the most effective immunorecognition capacity, as well as the best neutralizing capacity against the biological activities of venoms from
Bothrops spp.,
Bothrocophias spp.,
Bothriechis spp.,
Crotalus durissus,
Lachesis spp., and
Porthidium spp., followed by the Silanes (Bioclon) antivenom from Mexico
[190]. In a more recent study
[185],
B. asper venoms from Gorgona Island and the Pacific and Western ecoregions of Colombia were evaluated from immunoreactivity assays using INS and Probiol antivenoms. The results obtained indicate again that the INS antivenom had higher antibody titers and neutralization compared to Probiol antivenom, showing the greater neutralizing capacity for the venoms of
B. asper from Gorgona island population.
It is important to study the variability in the protein composition of different pitviper venoms and to develop studies that describe the structure-function correlations of individual toxins that will help to improve an understanding of venoms as trophic and defensive adaptations in different species
[213]. Likewise, it is important to continue exploring alternatives for the potential use of venom toxins as active compounds in the development of new drugs. However, this should be preceded by a deeper study of the natural history of viperid genera present in Colombia (e.g., distribution), to better understand how the destruction of their natural habitats and climate change affects the variability of their venoms and their survival in both natural and altered environments.
Appendix: Material and Methods
Phylogenetic analyses
Phylogenetic analyses were performed using maximum-likelihood estimation in the IQ tree software
[215]. The evolutionary models were performed using the model finder
[216]: TN+F+I: CYTB1_ND41, TIM+F+R2: CYTB2, GTR+F+I+G4: CYTB3_ND43, TN+F+R2: ND42. The analyses were performed using an array of 102 concatenated sequences for 51 terminals for the
Bothrops atrox complex and the
Porthidium genusfrom Colombia, Brazil, Ecuador, Venezuela, and Costa Rica. We used 20 species as out groups:
Gloydius halys,
Sistrurus miliarius and
S. catenatus;
Crotalus horridus,
C. ruber,
C. atrox, and
C. durissus;
Agkistrodon piscivorus and
A. contortrix;
Mixcoatlus melanurus;
Ophryacus ondulatus;
Cerrophidion godmani;
Metlapilcoatlus (Atropoides)
nummifer and
M. mexicanus;
Atropoides picadoi and
Bothrocophias hyoprora,
B. tulitoi and
B. myrringae;
Bothriechis schlegelii (currently redescribed as
Bothriechis nigroadspersus)and
Bothrops jararaca. Besides, phytools were used to perform a relationship analysis (heatmap) at the phylogenetic level
[217], using two published proteomes for Colombian pitviper species and some related lineages/clades.
Lethality test
The biological assays for lethality, coagulant, defibrinogenating, hemorrhagic, and edematogenic activity were conducted by the INS (National Institute of Health) as part of its antivenom production activities, in accordance with venom characterization guidelines established by the WHO in "Guidelines for the Production, Control and Regulation of Snake Antivenom Immunoglobulins; 2017". All tests were performed on CD1 strain mice weighing 18-22 g, following INS protocols for the use of laboratory animals.
Median Lethal Dose (LD50₅₀).— We assessed venom lethality following the procedures and protocols described by
[200,218], as well as the INS production protocol MEN-R04.6022-011. The median lethal dose (LD₅₀) of a venom is defined as the amount of venom that causes death in 50% of inoculated mice. To determine the LD₅₀, different doses of venom were prepared in saline solution and administered intraperitoneally to groups of five mice. Mortality was recorded after 48 hours. A control group of five mice received saline solution. Finally, the LD₅₀ was estimated using statistical methods such as the Spearman-Karber, Probit function, or other non-parametric methods
[219].
Minimum Coagulant Dose (MCD).— In vitro coagulation assays were performed following the procedures and protocols described by
[200,220], as well as the INS production protocol MEN-R04.6031-001. The minimum coagulant dose (MCD) was defined as the lowest venom dose that induced coagulation in citrated human plasma.
Minimum Defibrinogenating Dose (MDD).— In vivo defibrinogenation assays were conducted following the INS production protocol MEN-R04.6022-046, as well as the procedures and protocols described by
[200,220]. The MDD corresponds to the venom dose that, when administered intravenously to five mice injected with a venom dilution, induces incoagulability after 2 hours of sample collection.
Minimum Edematogenic Dose (MED).— In vivo edema assays were performed following the INS production protocol MEN-R04.6022-043, as well as the procedures and protocols described by
[200]. The MED is defined as the venom dose that induces a 30% increase in mouse paw edema following the injection of a venom dilution. For this purpose, venom solutions at different concentrations were prepared. Each dilution was injected into the right hind paw plantar pad of five mice, while the left paw received sterile saline (0.85%). One hour after inoculation, the animals were ethically euthanized
[221]. Both paws (at the tibiotarsal joint) were excised and weighed.
Minimum Hemorrhagic Dose (MHD).— In vivo hemorrhagic assays were conducted following the INS production protocol MEN-R04.6022-048, as well as the procedures and protocols described by
[200,220]. The minimum hemorrhagic dose (MHD) was defined as the smallest amount of venom capable of inducing a hemorrhagic lesion with a 10 mm diameter. To determine this, different venom dilutions in saline solution were prepared, with a final injection volume of 50 µL. Each dilution was then injected intradermally into the shaved ventral skin of five mice. After 2-3 hours, the animals were ethically euthanized
[221]. The skin was dissected, and the diameter of the resulting hemorrhagic lesion was measured.