1. A brief historical account of the snake species richness in Colombia
Historically, most of Colombian snakes have been poorly studied, producing significant gaps in our knowledge about essential aspects of their natural history traits, ecology, behavior, systematics, taxonomy, distributions, ophidism, ethnobiological uses, and social perceptions. Currently, some of this topics remain poorly understood
[1–3]. However, since the end of the 20th century and the first decade of the 21st century, a plethora of studies has emerged, aiming to reduce the historical deficit of understanding the snake species richness that inhabits one of the top megadiversity countries worldwide
[1].
The knowledge about the snakes of Colombia is limited for multiple reasons, such as: scanty funding resources for field studies; few holotypes (i.e., the specimen on which the description of a species is based) are housed in Colombian biological collections; most of the medical important snake species have few vouchers in Colombian biological collections, in turn representing limited geographic representativeness (see below); few experts in universities and research institutions are capable of describing and identifying snakes, and teaching about their biology; vast areas of the country, as well as several type localities (i.e., the locality where the holotype of a species was collected) of most Colombian venomous snakes, have been historically inaccessible or highly risky for scientist due internal armed conflicts
[4–6].
Because of the above, large regions of Colombia still lack intensive sampling, and most of the Colombian ophidiofauna has not been subject to comprehensive biological studies. As a result, significant sampling and knowledge gaps exist, rendering Colombia a “black hole” in terms of essential biological knowledge for most groups of medically significant snake species, despite its key geographical position in the historical exchange of faunas between North and South America
[7], as well as being one of the countries with the highest annual rate of reported snakebites in America (see Chapter 9).
As a member of the Royal Botanical Expedition, and zoological components manager, Jorge Tadeo Lozano
[8,9], was the first Colombian to study Colombian amphibians and reptiles. He particularly emphasized snake natural history, and the non-medical practices used to deal with snakebites during the end of the period of the Nuevo Reino de Granada (1538-1819) and the beginning of the Gran Colombia period (1819-1831). Thus, he became the first Colombian herpetologist
[9] (see Chapter 7). Particularly, Jorge Tadeo Lozano in his publication titled “Memorias sobre serpientes”stated future research guidelines that would increase our understanding of snakebite and its eco-epidemiological context in Colombia (see Chapter 9), as follow
[10]:
- To study the snake anatomy for classifying venomous from non-venomous snakes, evaluating the oral structures very carefully to ensure the existence of "venom accumulator bags (=venom glands)" and hollow fangs (=solenoglyphus dentition).
- To study the activity and effect of snake venoms, using animals of varied sizes and types, scrupulously noting all the phenomena that arise.
- Once the effects of each venom are known, test whether they can be used in medicinal treatments for diseases, or to combat the venoms of other species.
- Evaluate the effect of removing the venom apparatus from snakes.
- Examine the amount and intensity of snake venoms, depending on their age, sex, size, times of year, and other circumstances.
- Perform a chemical analysis of the "venom liquor", recognizing its nature, composition, and differences between species.
- Study the "anti-venoms" that are commonly used, and test their effectiveness on bitten animals, or by mixing them with venoms, and observe if they decompose, disorganize, or have deprived them of their deleterious virtue.
- Do all experiments for evaluating the guaco plant (=
Mikania guaco), the most used "anti-venom."
- To investigate the amount of venom required to kill an animal of each order.
- Make a detailed study of venomous snakes to distinguish them from harmless ones, using the characteristics of number, arrangement and figures that make up their scales.
- To find out if nature gave the snakes venom solely for self-defense and to kill their enemies, or if, as I suspect, it is a supplement to aid in digesting food instead of chewing it, which they cannot do due to the structure of their teeth that prevent it.
Thew Lozano's guidelines were visionary because most of his recommendations and conjectures are the currently most important and prioritized lines of research of venomous snakes, as well as dealing with snakebite envenoming
[11–13] (see Chapter
3,
5,
6,7,9,and 10). After Lozano's visionary guidelines, national and foreign researchers attempted to describe the natural history and envenomation capacities of the snakes inhabiting the region at the beginning of the birth of the country now known as Colombia. Most of these researchers were motivated to provide an understanding of snakebite accidents and the therapies used as medical practices.
In 1870, the engineer and homeopathic doctor Silvestre B. Higgins, native of the city of Barranquilla (Atlántico department), published the book “Culebrasi reptiles venenosos: sus hábitoscaracterísticas i particularidades”. This book focuses on the non-medical practices used as therapies against snakebites by Colombian healers. It also describes general aspects of the natural history traits of some venomous species and its ecoepidemiological context. He compiled the most amazing beliefs, tales, myths, and magical events regarding Colombian snakes, explaining their common names, and social beliefs that still prevail in many rural communities.
Interestingly, Higgins provided an annotated list of venomous snakes from each of the nine Colombian states (Higgins lived during a history period in which Colombia was a cluster of federal states named as Estados Unidos de Colombia 1863-1886). However, Higgins did not provide a solid scientific foundation in his book, so venomous and non-venomous snake species were listed by common names rather than scientific ones. Thus, based on Higgins’s species list, the results are difficult to apply reliably to most of the snake taxa described. However, by excluding common name synonyms, non-snake species (e.g., amphisbaenians), and non-venomous species (e.g.,
Pseudoboa neuwiedii) from Higgins’s list, the number of snake species can be estimated to represent 12:
Bothrops asper,
B. atrox,
B. bilineatus,
B. punctatus,
Bothrocophias spp.,
Bothriechis schlegelii,Crotalus durissus,
Micrurus dumerillii,
M. mipartitus,
Lachesis acrochorda,
L. muta, and
Porthidium lansbergii.
In 1889, the physician Andrés Posada Arango published the article “Note Sur Quelques Solenoglyphes de Colombie” in which he describes four new species from the antiquated genus
Thanatophis (Thanatophis patoquilla, T. sutus, T. montanus, and T. torvus) according to his interpretation. Nevertheless, these species were poorly described and posteriorly synonymized into three genera and three different species or species complexes (T. patoquilla and
T. sutus were synonymized with
Porthidium lansbergii[14];
T. montanus was synonymized with
Bothrops punctatus[15],and
T. torvus was allocated to the genus
Bothriechis as
B. torvus[16]).
.png)
Figure 1. Illustrations of the new species proposed by Andrés Posada-Arango. (Top) Head in lateral view of
Bothriechis schlegelii species complex from Manizales, Caldas (INSV-SR-00138). (Bottom) Head in lateral view of
Porthidium lansbergii (INSV-SR-89) from Yondó, Antioquia, Colombia. Illustrations by Oscar Ramírez.
In 1896 the physician Evaristo Garcia Piedraita provided a list of medically important snake species from the Cauca department (currently Cauca, Valle del Cauca, and Putumayo departments). Dr. Garcia-Piedraita in his book “Serpientes venenosas del Cauca”[17], provides a detailed study of the classification of venomous snakes; and he characterizes, describes, and diagnoses the venomous apparatus, the effects of venoms, and the symptoms of envenomation caused by snakebites. Additionally, he analyses and reflects on the empirical methods used against snakebites, methods to neutralize venom, and the treatment to counteract snakebite symptoms
[1]. Garcia-Piedrahita provided a list and description of 19 medically important snake species, however, discounting common name, synonyms, and non-snake species and non-venomous species from the Garcia-Piedraita’s species list, a total of 11 species were reported (Bothriechis schlegelii,
Bothrops asper,
B. rhombeatus,
B. punctatus,
Crotalus durissus,
Lachesis acrochorda,
L. muta,
Porthidium lansbergii,
Micrurus mipartitus, and
M. hemprichii).
Two decades later, Afranio do Amaral
[18,19] expanded the knowledge about Colombian snakes providing a regional species list of snakes of Colombia, reporting five venomous snakes from the Santa Marta region (Micrurus dumerillii,
M. mipartitus,
Bothrops asper,
Crotalus durissus,
Porthidium lansbergii,) and six from the San Juan River lower Basin, Chocó department (Micrurus ancoralis,
M. mipartitus,
Bothrops asper,
B. punctatus,
Bothriechis schlegelii,
Porthidium nasutum). Nevertheless, until the 1940s, Nicéforo María
[20] was the first researcher who attempted to consolidate a complete species account list of Colombian snakes. He reported about 180 snake species, however, discounting subspecies from Nicéforo’s species, his list the number of snake species reaches 174 (pers. obs. J.D. Lynch)
[1]. A total of 26 venomous snakes were reported of which 15 were elapids (with the description of
Micrurus sangilensis as a new species), and 11 viperids.
In 1968, Federico Medem (Friedrich Johann Graf von Medem)
[21] provided a comprehensive review of the historical development of herpetology in Colombia, listing all known species of amphibians and reptiles of the country, encompassing records from explorers from XVIII century up to 1968. He reported a total of 231 snake species inhabiting Colombia, of which, discounting subspecies, 10 were viperids and 13 were elapids. Medem provided detailed descriptions of their geographic distributions, habitats, natural history traits, ethnozoological data, and social beliefs.
Two years later, Peters and Orejas-Miranda
[15] provided a comprehensive catalogue of the Neotropical snakes, including taxonomic keys for genera, species, and subspecies. They provided scientific names, distributions and attempted to resolve unknown or undefined taxa. Discarding subspecies, these authors reported 182 Colombian snake species of which 16 were viperids and 17 elapids. The taxonomic keys for genera, species, and subspecies remain valid for many taxa, and these continue to be used in several biology academies institutions in Latin America as an introductory approach to an understanding the taxonomic richness of Neotropical snakes.
In 1985, a new attempt to compile knowledge about Colombian venomous snakes was performed by the physician and professor of the Universidad de Antioquia, Rodrigo Ángel
[22] when he published the monograph “Serpientes de Colombia: guía práctica para la clasificación y tratamiento del envenenamiento causado por sus mordeduras”. Ángel reported 160 snake species for Colombia of which 13 were viperids and 31 elapids. However, discounting subspecies from Ángel’s species list the number of venomous snakes in Colombia reaches 36 taxa (Figure 2). This book was one of the main references for most of the medical personnel who faced snakebites in Colombia up to the end of the 20th century.
In 1988, Pérez-Santos and Moreno published the first modern monograph, “Ofidios de Colombia”, that extensively compiled the snake species richness of Colombia. They provided keys for their identification, descriptions of their diagnostic characters, and maps of their distributions. These authors reported 240 snake species in Colombia discounting subspecies. A total of 36 venomous species were reported (20 elapids and 16 viperids). Nevertheless, this monograph has significant shortcomings in the quality of species records, mainly because most of the distribution records and natural history information is inadequately documented, presenting questionable identifications and distributions, totally lacks cross-references of the specimens per species and museum collection numbers
[23]. Despite this, the book provided comprehensive species checklists and diagnostic characters, most of it useful. It lacks a rigorous evaluation of the data used, resulting in an underestimation of species richness for the country, and adds confusion regarding their distributions within the national territory
[1,23].
In 1989, Campbell and Lamar offered a comprehensive and well-supported account of venomous snake species across Latin America, including Colombia. Their work addressed several misconceptions raised by previous publications and corrected data of previous studies that often-lacked cross-references of specimens per species and museum collection numbers, or else omitted references from published data, resulting in confusing geographic distributional ranges.
Towards the end of the 20th century, Sanchez et al
[24] provided a biological review of reptile species in Colombia
[24]. However, in this publication, the quality of the records used was not evaluated. This oversight led to the an underestimation of species richness for the country and confusion regarding their distributions within the national territory.[1]. This resulted in an underestimated species richness for the country (Figure 2).
Since the 21st century an explosion of studies has emerged, aiming to reduce the historical deficit in understanding the snake’s richness. These studies have quickly increased the number of snake species with comprehensive taxonomic reviews, as well as given the description of new species, thus, passing from ~240 to ~331 snake species (Figure 2). In 2002, Pineda and Renjifo presented and updated account of Colombian snakebite accidents, focusing on clinical reports, effects of venoms, and the symptoms of envenomation caused by snakebites present in Colombia, providing a comprehensive overview of the clinical manifestations, epidemiological information, and how to handle snakebite accidents and their therapy and updating that previously presented by Ángel in 1985. Also, these authors provided an account of 35 venomous snake species. However, they cited inaccurately the venomous snake list previously provided by Campbell and Lamar
[25], missing some
Micrurus species (Figure 2).
.png)
Figure 2. Overview illustrating the exponential growth of snake richness in Colombia. The size of the bubbles corresponds to the ratio of venomous snakes to total snake species, as reported by each author. The number linked with each bubble indicates the number of venomous snakes reported by that author.
In 2004, Campbell and Lamar
[26], made notable advances toward under-standing venomous reptiles in the Western Hemisphere. The classic book “The Venomous Reptiles of the Western Hemisphere” is the first modern compendium that setting the modern baseline for understanding the taxonomic conundrums of Colombian venomous snakes. This comprehensive study updated their proposal from 1989 and add significant information. Their taxonomic proposals remain valid for many species of venomous snakes distributed in Colombia.
In 2016, Cañas et al.
[27], in their book “Serpientes venenosas: lecciones aprendidas desde Colombia”, provide an interesting review of the knowledge accumulated until the 20th century and the beginning of the 21st century on the diversity, general biology, and identifying characteristics of medically significant snakes in Colombia. They describe the features that allow for distinguishing venomous snakes from non-venomous ones and offer a clinical perspective on snakebite manifestations. They are the first authors to compile toxinological advances related to venomics, biochemistry, and the biological activities of venoms from the medically important snakes in the country. Additionally, they presented epidemiological information and guidelines for managing snakebite, with emphasis on snakes distributed across the four life zones of the Valle del Cauca department. These authors indicate that 310 snake species inhabit Colombia, 53 are medically important snakes (21 vipers and 32 elapids), of which 14 species are distributed in Valle del Cauca department. However, their list of species lacks cross-references with specimens from collections or published literature, making it impossible to trace the data's sources.
In 2021, Ayerbe
[28], in his book “Serpientes: accidentes y soluciones” provides a detailed description of the symptoms and signs of poisoning caused by Colombian snakes, their clinical manifestations and complications in their treatment, presenting therapeutic approaches to deal with the snake accident. Within the generalities of the biology of snakes of medical importance in Colombia, Ayerbe
[28] indicates that 310 species of snakes live in the country, of which 56 are of medical importance. However, similarly to Cañas et al.
[27], he does not provide cross-references with specimens from collections or published literature to support his lists (e.g.,
Micrurus hemprichii versus
Micrurus ortoni; see the explanation in the following sections of this chapter).
Since Campbell and Lamar
[26], most studies of the 21st century have corrected the shortcomings of the past, providing snake accounts based on high-quality records from peer-reviewed literature, museum collection specimens, and curated digital species repositories or databases
[1]. The ongoing project of Uetz, Hallermann, Hosek, and collaborators, the Reptile Database (http://www.reptile-database.org), have accelerated and consolidated the historical efforts previously undertaken by several authors, providing a modern method to annually update species lists of worldwide non-avian reptile species.
Currently, this website serves as the primary or initial option for any researcher or reptile enthusiast seeking a basic understanding of taxonomic richness, geographic distributions, specialized literature, original descriptions, pictures, and other relevant biological data. The Reptile Database representing the “modern consensus” among herpetologists worldwide about scientific name, taxonomic decisions, and distribution. No doubt, Uetz, Hallermann, Hosek et al. have made an enormous advance in compiling a colossal amount of knowledge about non-avian reptile species
[29].
Nevertheless, since the Reptile Database do not provide any taxonomic assessment beyond a checklist with the feedback from some the authors who proposed it; and these species list is based on information from various sources
[30], controversy about taxonomic decisions and distributions of several taxa, especially snakes, persists. As taxonomy is a dynamic scientific field, controversy always fuels its engine. Thus, the total number of snake species could change between authors or within the same authors across time according to the criteria employed by them, generating a constant expansion and contractions of the snake species list due to redundancy or missing taxa.
For example, in this book, we consider that currently in Colombia there are a total of 331 snake species, of which 49 taxa represent venomous snake species (Viperidae and Elapidae; see
https://ofidismo.ins.gov.co). Our species accounts varies from Uetz et al
[29] for 4.6% of the total snake species and 18% for venomous species account (Figure 2, Table 1). The criteria employed that explain these differences are included in the following sections of this chapter. Despite the variability in snake species lists among authors, in the 21st century, Colombia has gained an exponential understanding of its ophidiofauna, indicating that the country has begun to emerge from the "black hole" by acquiring essential snake biological knowledge. Across time, the ratio of venomous snakes to total snake species has decreased, indicating that the medical view of snakes gradually changed towards a broader biological view including all snakes in the species account list.
Furthermore, since the beginning of the 21st century this ratio has had averaged of 16.3% (9.8-20.1%) indicating that the medically important snake species in Colombia represented less than 20% of the total ophidiofauna of the country. We hope that soon, this trend continues by addressing the taxonomic uncertainties of several Colombian snake taxa. However, this historical trend also points out that our understanding of Colombian snakes is far from comprehensive, encouraging us to continue investing significant efforts due to their crypticity and the dearth of information surrounding them. We hope this challenge will be addressed by both young national and international herpetologists.
2. A megadiverse mess: The taxonomic conundrum of the medically important snake species in Colombia
One of the key stages for any person or medical practitioner facing a snakebite accident is the accurate identification of snake. According to the snake taxonomic identification, medical practitioner can provide an accurate report to the health surveillance system, to determinate the envenoming type, addressing the suitable treatment and antivenom therapy, as well as anticipating the possible clinical complications
[31]. For this reason, most of the guidelines, manuals, field guides and protocols aiming to treat or manage snakebite accidents, include the snake taxonomic identification as a paramount stage into its step-by-step process (algorithm)
[27,28,31–34](see Chapter 9).
Despite this task sounding clear and easy, in tropical and megadiverse country like Colombia the task can be complicated, even more when several of the snake species have a puzzling taxonomic status due to poor scientific research and their high crypticity
[1]. Additionally, the distributional of several venomous snake species can extensively overlap (sympatric species), rendering species identification through the geographical elimination method unsuitable in most cases.
Recently, several researchers have made important efforts to untangle the taxonomic problems of some Colombian snakes of medical importance
[35–38]. Nevertheless, most snake diversity in the country continuous with puzzling taxonomic status, maintaining as a taxonomic baseline the classic and comprehensive taxonomic reviews made in the end of the 20th the beginning of the 21th centuries
[15,23,39–44].
Most of these scientific publications conclude that Colombian populations of venomous snakes require detailed taxonomic revision
[15,26,39,45–47]. Despite this, several researchers have employed synonyms and subspecific epithets as full species names without a taxonomic revision, or they lack solid evidence for the use, erection, or resurrection of a name
[28,48]. Hence, this practice has caused a taxonomic instability, deepening the conundrum of Colombian snakes of medical importance, beyond the science-based changes in taxon names
[49].
This phenomenon has occurred not only with Colombian snakes. Globally, taxonomic issues arise when the data used to create taxonomic decisions are shoddily presented, derive from spurious research, or lack evidence
[50]. Thus, subsequent application to a broad range of endeavors, like conservation or neglected tropical diseases (e.g., ophidism), undermine the results by a lack of consistency. In addition, due to the explosion of electronic information and the social media revolution across of the first two decades of 21th century
[51], a rapid dispersion and merge of scientific and non-scientific taxonomic information has occurred, making it difficult to make suitable taxonomic decisions for medical practitioners or non-herpetology professionals.
Accordingly, Kaiser et al.
[50] propose that a scientific taxonomic decision must follow three main steps: (1) generate hypotheses of group membership (e.g., a species, a clade or taxon) or evolutionary relationship (e.g., sister taxa) based on available primary sources (e.g., fossil record, existing or new collections of specimens including whole animals, tissues, quantitative analyses, and DNA sequences, etc.) and the available literature; (2) test these hypotheses via appropriate, rigorous, and honest analysis of the relevant data using the scientific method; and (3) submit proposed taxonomic decisions (e.g., taxonomic rearrangements, descriptions of new species, elevation of subspecies to species rank) to peer-reviewed journals in the form of manuscripts that present the data and provide a rational justification for the proposed decisions.
Unfortunately, several high impact investigations did not follow the Kaiser et al.
[50] approach, instead basing taxonomic decisions on lacking evidence, evidence shoddily presented; or worse, committing taxonomic vandalism
[52,53], that means the deliberate establishment of scientific names (= hypothesis) by eschewing the scientific process described above
[49,50]. In addition to these shortfalls, some taxonomic decisions have been published in predatory/fake journals or fast-track journals, which in exchange for a payment, the authors get a rapid publication and broad audience, at the expense of scientific quality, legitimacy of the taxonomic decisions, and the robustness of the peer-review process, and scientific value.
Confusion regarding species or genus names of venomous snakes could cause serious constraints in the implementation of public health schemes and snakebite treatment. For example, this can increase the underreporting of snakebite cases by classifying them under a species name that is nonscience-based, as well as masking the variability of clinical symptoms of envenoming from broadly distributed species
[54–57].
Particularly in Colombia, one of the main difficulties during the diagnostic and snakebite accident treatment is the correct snake species or genus identification
[1]. This arises as one of the main obstacles constraining the need for quick decision-making by a medical practitioner for the suitable antivenom therapy required, including the antivenom dosage, as well as the prevention of clinical complications during the treatment
[58].
The species concept continues to be debated due to the fact that no single species concept simultaneously include all the ways, forms and combinations in which life evolves
[59]. Nevertheless, all species concepts have something in common: all consider species as a hypothesis. Hence, all taxa can be treated as “the educated assumption of some outcome based on scientific method, logic and observation”
[60]. Thus, all can be the subject of testing. To treat species as hypotheses allows employing multiple lines of evidence as operational criteria (sensu Mayden
[61]), to perform empirical testing and diagnosis of species in the nature
[61–63].
According to this outlook, in this section we present a reviewed venomous species list based on the currently available information regarding the taxonomic status of the medically important snake species, encompassing colubrids (aglyphous and opisthoglyphous) viperids (solenoglyphous), and elapids (proteroglyphous) inhabiting Colombia. We conducted a detailed review based on the available literature in order to make the “best” scientific taxonomic decisions following the Kaiser et al.
[50] proposal, as well as all those that clustered around the major lines of evidence to delimiting a species.
Our aim is not to resolve all current taxonomic problems of Colombian snakes, but to provide taxonomic treatment and criteria to allocate the puzzling snake species into a taxonomic scheme based on the current evidence available. This will help to medical to designate the taxonomic snake entities that cause bites in Colombia, reducing misunderstanding and misidentification, as well as the underreporting rate of snakebite
[56,64] (see Chapter 9).
We hope that the next taxonomic review based on a scientifically based approach (including integrative taxonomy) and publication in a high quality peer-reviewed journal can untangle the puzzling Colombian snake species included here. All the criteria defined in this chapter will include the species taxonomic treatment throughout the chapters of this book.
2.1 Tackling the taxonomic uncertainties in the medically important snake species
During the last two decades, with the explosion of molecular techniques and analyses, the massive use of quantitative analyses, bioinformatic, high resolution computed tomographies (HRCT) aided by available free open-source software available (e.g., R, Python, Past, etc.), and the popularization of the hemipenial eversion techniquesamong researchers
[65], new species, genera resurrection, revalidation or elevation of species from subspecies, and species synonymies have been proposed
[35,36,66–71]. These efforts, most of them including multiple lines of evidence (integrative taxonomy), have allowed us to untangle several taxonomic problems of South America venomous snakes previously pointed out by Campbell and Lamar
[26], providing a notable advances toward our understanding.
Nevertheless, in comparison with these efforts,few taxonomic attempts using multiple lines of evidence have been made to resolve some puzzling Colombian snake venomous species
[35,37,72]. In most Colombian species Campbell and Lamar
[26] continues to be the paramount guideline for taxonomic decisions because there is no new evidence to tackle problematic species (e.g.,
Micrurus dumerilli,
M. mipartitus,
Bothrops venezulensis), or the new evidence provided has been shoddily presented, causing more confusion than clarity (e.g.,
Bothrops ayerbei and
Bothrops rhombeatus;
see Chapter 3).
Taxonomic problems arise from multifactorial causes. However, we summarize them in two main categories as follows:
Crypticity.— Researchers can observe insights indicating that populations of a particular species could represent different evolutionary lineages, but under the current state of knowledge, they cannot readily be distinguished or delimited due to significant uncertainties of their diagnostic characters (e.g., indistinct morphological attributes and/or ecological properties, lack of DNA data, or available quantitative analyses available)
[63,73,74]. Therefore, a taxon might be considered as a complex of species that could cluster two or more species, but new lines of evidence are required for a suitable diagnosis and delimitation
[74].
Data deficient.— This category groups species that: (1) since its erection and type series (the group of specimens on which the description of a species is based in addition to the holotype), no new specimens or populations are known; (2) lack or loss of type specimens; (3) species erected from a single known specimen; (4) its taxonomic name cannot be assigned with certainty to any taxonomic group because the description is insufficient for identification, and/or the original specimen is lost, or no longer exists (nomen dubium)
[75]; (5) despite having been the object of a taxonomic revision across its known distribution (employing integrative taxonomic approach or a single line of evidence), Colombian populations of this species have not been included in these assessments; and (6) species delimitation and diagnosis has deep shortfalls and incongruences during its establishment. Therefore, conclusions made about its taxonomic status cannot be straightforwardly assumed for Colombian populations.
Using a taxonomic conservative view, the Colombian medically important snake species list is proposed here (Table 1), is based on a comprehensive revision of the available lines of evidence for the species delimitations, as detailed in the following section.
2.2 Records of Colombian medically important snake species
We retrieved and curated approximately 5,488 records of medically important snake species geolocated in Colombia from 50 sources. These references include specimens housed in biological collections, records from specialized herpetological literature (articles, books), unpublished data from field notebooks of various Colombian and foreign herpetologists, and clinical records from Colombian serpentarium dedicated to antivenom research and production (see
https://ofidismo.ins.gov.co).
Colombia harbors a total of 59 medically important snake species, of which ten species are colubrid, 20 are viperids, and 29 are elapids. About 13.6% of these species bite frequently and are associated with serious and life-threatening envenoming, while the remainder of snake species rarely causes serious and life-threatening envenoming or have no reported cases (Table 1).
Table 1. Colombian medically important snake species. m: meters above sea level.
| Species |
Endemic |
Type locality in Colombia |
Department Distribution |
Elevation range (m) |
| Snakes that bite frequently associated with serious and life-threatening envenoming |
|
Bothriechis schlegelii* |
No |
Yes |
Ant, Boy, Cal, Cau, Ch, Cor, Cu, Hui, Nar, Qui, Ris, San, Tol, Val |
2—2946; = 1832 |
|
Bothrops asper* |
No |
No |
Ant, Atl, Bol, Boy, Cal, Cau, Ces, Cho, Cór, Cun, LaG, Mag, NSa, Qui, Ris, San, Suc, Tol, Val |
2—2200; = 547 |
|
Bothrops atrox* |
No |
No |
Ama, Arau, Boy, Caq, Cas, Cun, Gua, Guav, Met, NSa, Put, Vau, Vic |
57—1923; = 308 |
|
Crotalus durissus |
No |
No |
Ant, Ara, Atl, Bol, Boy, Ces, Cun, Hui, LaG, Mag, Met, Tol, Vic |
7—1717, = 235 |
|
Porthidium lansbergii* |
No |
Yes |
Ant, Atla, Bol, Boy, Cas, Ces, Cho, Cór, Hui, LaG, Mag, NSa, San, Suc, Tol |
0—1243, = 782 |
|
Porthidium nasutum |
No |
No |
Ant, Cho, Val |
12—1011, = 307 |
|
Micrurus dumerilii* |
No |
Yes |
Ant, Atl, Bol, Boy, Cal, Cau, Cesar, Cho, Cór, Cun, Hui, LaG, Mag, Met, Nar, NSa, Ris, San, Suc, Tol, Val, |
2—2278, = 815 |
|
Micrurus mipartitus* |
No |
Yes |
Ant, Boy, Cal, Cau, Ces, Cho, Cór, Cun, Hui, Mag, Met, Nar, NSa, Ris, San, Suc, Tol, Val, |
11—2606, = 1204 |
| Snakes that bite frequently, but rarely cause serious and life-threatening envenoming |
|
Dryophylax gambotensis |
Yes |
Yes |
Atl, Bol, Ces, Cór, LaG, Mag, Suc |
1—161, = 34 |
|
Helicops angulatus |
No |
No |
Ama, Caq, Cun, Gua, Guav, Met, Put, Vau, Vic |
52—570, = 237 |
|
Leptodeira annulata* |
No |
No |
Ama, Ant, Atla, Bol, Boy, Cal, Caq, Cas, Cau, Ces, Cho, Cór, Cun, Hui, LaG, Mag, Met, San, NSa, Suc, Tol, Val, Vic |
3—2130, = 646 |
|
Oxybelis fulgidus |
No |
No |
Ama, Bol, Cat, Mag, Met, Suc, Vau |
3—484, = 220 |
|
Erythrolamprus bizona* |
No |
Yes |
Ant, Boy, Cal Cas, Cau, Ces Cun, Hui, Mag, Met, NSa, San, Tol, Val |
14—2566, = 900 |
| Thamnodynastes pallidus |
No |
No |
Ama, Cas, Met, Put, Vic |
75—437, = 162 |
|
Xenodon rabdocephalus |
No |
No |
Ant, Cal, Cau, Cho, LaG, San, Tol, Val |
3—1778, = 483 |
| Snakes that bite rarely, but are capable of causing serious and life-threatening envenoming |
|
Bothrocophias myersi |
Yes |
Yes |
Cau, Cho, Val |
12—1275, = 224 |
|
Bothrocophias myrringae |
Yes |
Yes |
Cun, Met |
1757—2761, = 2137 |
|
Bothrocophias tulitoi |
Yes |
Yes |
Boy, Cas, Cun |
1685—2694, = 1969 |
|
Bothrops punctatus |
No |
Yes |
Ant, Cal Cau, Cho, Nar, Ris,Val |
4—1578, = 569 |
|
Hydrophis platurus |
No |
No |
Cau, Cho, Nar |
-50—0, = -5 |
|
Lachesis acrochorda |
No |
Yes |
Ant, Boy, Cau, Cho, Nar, San, Val |
2—1775, = 655 |
|
Lachesis muta |
No |
No |
Ama, Caq, Met, Put, Vau |
24—1809, = 352 |
|
Micrurus helleri |
No |
No |
Ara, Caq, Cas, Cun, Met, Put, Vau |
87—1338, = 393 |
|
Micrurus hemprichii* |
No |
Yes |
Ama, Boy, Caq, Cas, Met, Vic |
67—770, = 311 |
|
Micrurus lemniscatus* |
No |
No |
Ama, Gua |
79—125, = 84 |
|
Micrurus obscurus |
No |
No |
Ama, Caq, Gua, Guav, Met, Put, Vau, Vic |
79—616, = 243 |
|
Micrurus surinamensis |
No |
No |
Ama, Caq, Cun, Guav, Met, Vau, Vic |
52—537, = 292 |
| Snakes that bite rarely, and have not caused significant envenoming or have not caused documented bites |
|
Bothrocophias campbelli |
No |
No |
Cho, Nar, Val |
32—1650, = 428 |
|
Bothrocophias colombianus |
Yes |
Yes |
Cho, Cau, Ris |
211—2506, = 1530 |
|
Bothrocophias hyoprora |
No |
No |
Ama, Caq, Guav, Nar, Put, Vau |
64—1202, = 234 |
|
Bothrops bilineatus |
No |
No |
Ama, Caq, Vau, |
80—265, = 105 |
|
Bothrops oligobalius |
No |
No |
Ama, Caq, Guav, Vau |
81—274, = 151 |
|
Bothrops pulcher |
No |
No |
Caq, Put |
274—1725, = 872 |
|
Bothrops taeniatus |
No |
No |
Ama, Cun, Gua, Met, Vau, Vic |
78—533, = 155 |
|
Bothrops venezuelensis |
No |
No |
Boy, Cas |
151—1785, = 1052 |
|
Micrurus ancoralis |
No |
Yes |
Ant, Cal, Cho, Ris, Tol, Val, |
21—707, = 98 |
|
Micrurus camilae |
Yes |
Yes |
Ant, Cór, San, Suc, |
30—1381, = 178 |
|
Micrurus clarki |
No |
No |
Cau, Cho, Val |
2—865, = 257 |
|
Micrurus dissoleucus |
No |
No |
Atl, Bol, Ces, Cór, LaG, Mag, NSa, Suc, |
5—1297, = 124 |
|
Micrurus filiformis |
No |
No |
Ama, Caq, Cas, Gua, Guav, Met, Vau, Vic |
68—438, = 205 |
|
Micrurus isozonus |
No |
No |
Ara, Cas, Met, Vic |
4—429, = 276 |
|
Micrurus langsdorffi |
No |
No |
Ama, Caq, Gua, Guav, Met, Put, Vau, |
81—637, = 179 |
|
Micrurus medemi |
Yes |
Yes |
Cun, Met |
229—1599, = 490 |
|
Micrurus multiscutatus |
No |
Yes |
Cau, Cho, Ris, Val |
67—2506, = 425 |
|
Micrurus narduccii |
No |
No |
Ama, Caq, Gua, Put, Vau |
75—283, = 140 |
|
Micrurus nattereri |
No |
No |
Gua, Vau |
85—229, = 166 |
|
Micrurus nigrocinctus |
No |
No |
Ant |
6—32, = 18 |
|
Micrurus oligonaellatus |
Yes |
Yes |
Cau |
1442 |
|
Micrurus ornatissimus |
No |
No |
Ama, Vau |
82—115, = 93 |
|
Micrurus psyches |
No |
No |
Ara, Met |
156—452, = 299 |
|
Micrurus remotus |
No |
No |
Gua, Vau |
83—233, = 128 |
|
Micrurus renjifoi |
Yes |
Yes |
Vic |
115 |
|
Micrurus sangilensis |
Yes |
Yes |
Boy, Cas, San |
993—2420, = 1660 |
|
Micrurus scutiventris |
No |
No |
Ama |
79—126, = 114 |
|
Micrurus spurrelli |
Yes |
Yes |
Cho |
41—71, = 51 |
|
Micrurus tikuna |
No |
No |
Ama |
78—81, = 79 |
| Other potentially medically important which have not caused documented bites |
|
Erythrolamprus aesculapii |
No |
No |
Ama, Caq, Guav, Put, Vau |
88—327, = 169 |
|
Leptophis ahaetulla* |
No |
No |
Ant, Cas, Ces Cun, Hui, Met, San, Tol, |
42—2171, = 746 |
Colombian departments. Ama: Amazonas; Ant: Antioquia; Arc: Archipiélago de San An-drés, Providencia and Santa Catalina; Arau: Arauca; Bol: Bolívar; Boy: Boyacá; Cal: Caldas; Caq: Caquetá; Cas: Casanare; Cau: Cauca; Ces: Cesar; Cho: Chocó; Cór: Córdoba; Cun: Cu-ndinamarca; Gua: Guainía; Guav: Guaviare; Hui: Huila; LaG: La Guajira; Mag: Magdalena; Met: Meta; Nar: Nariño; NSa: Norte de Santander; Put: Putumayo; Qui: Quindío; Ris. Risaralda; San: Santander; Suc: Sucre; Tol: Tolima; Val: Valle del Cauca; Vau: Vaupés; Vic: Vichada.
* = species complex.
The ten Colombian departments with the greatest snake species richness are Amazonas, Vichada, Vaupés, Chocó, Cauca, Caquetá, Antioquia, Cundinamarca and Valle del Cauca, ranging from 15 to 22 snake medically important species (Figure 3A). All departments encompass the most diverse ecoregions and complex topographic areas of Colombia and northern South America. However, most records are concentrated in the departments of Antioquia, Meta, and Santander because there are the political-administrative divisions of Colombia that historically have received major sampling efforts
[1].
When snake richness was analyzed after splitting into elapid and viperid species, the general tendency observed was stable, and the species richness clustered over the complex topography and ecologically diverse regions. However, for viperids, the departments with the greatest snake species richness were those located in the trans-Andean region of Colombian (Chocó, Valle del Cauca, Boyacá, Antioquia), while for elapids, species richness is mostly concentrated in the cis-Andean region (Meta, Amazonas Vichada, Vaupés, Caquetá; Figure 3B). This suggests intricate biographic patterns of expansion and diversification in the lineages of these venomous snake species (see Chapters
2 and
3). Antioquia, Arauca, and Meta are the departments most of the known geographic records of viperids clustered, while for elapid species, Santander, Cundinamarca, and Valle del Cauca were the departments that harbor most of the known geographic records (Figure 3C).
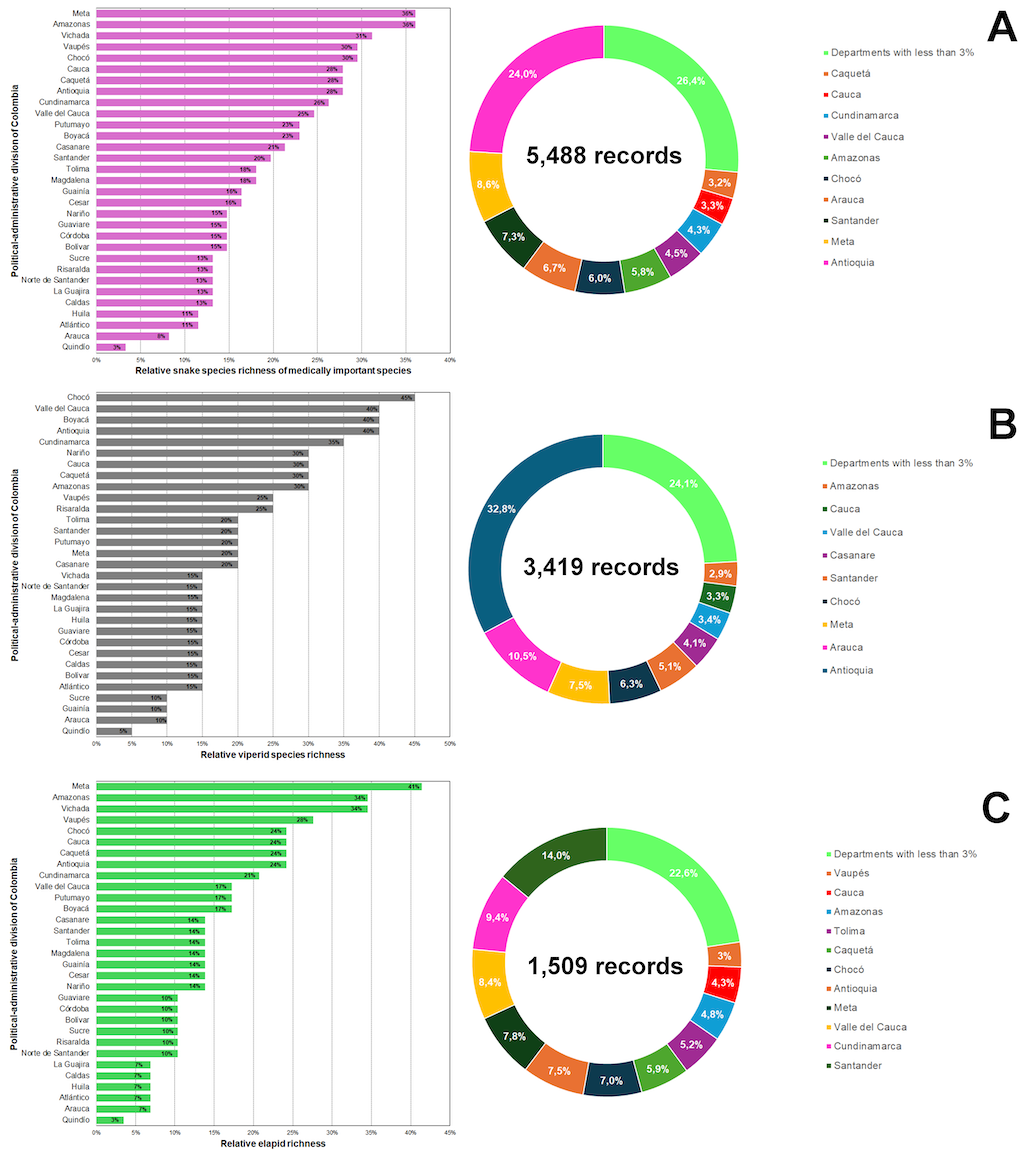
Figure 3. Species richness and records of medically important snake species of Colombia
Since political-administrative divisions are not biological constraints for species distributions, nor are they a suitable approach for estimating the geographical representativeness of medically important snake species, we divided the country into grids/pixels of 1.0 geographical degrees (110 x 110 km; WGS84 system) to analyze distribution. This approach allows for comparisons with previous studies, such as Lynch et al,
[1], and future research (Figure 4).
Regardless of the taxonomic group (viperids or elapids), most of the available records come from the trans-Andean region of Colombia, highlighting a notable historical disparity in the sampling effort to understand and document the medically important snake species in the country (Figure 4A-C). Additionally, this disparity is exacerbated when comparing viperids and elapids, showing that elapids have been historically under sampled (Figure 4D).
The greatest number of historical records of venomous snakes from the Viperidae and Elapidae families is concentrated in the department of Antioquia. This department also shows the highest annual average of snakebite reports for the period 2010-2020 (see Chapter 9), indicating that it is the only department that has the most robust information on two of the most important variables for understanding snakebite as an epidemiological event
[64]. In contrast, the rest of the national territory exhibits a notable under sampling of venomous snakes, with fewer than 112 records per pixel (110 km x 110 km; see Figure 4A). This represents a critical problem for understanding the total diversity of medically important snakes involved in human-snake conflict, as well as their incidence in snakebite events.
This knowledge gap exacerbates the limitations in understanding the causes of negative encounters between humans and snakes, identifying the species involved, and formulating and implementing strategies to prevent envenoming and conserve the country's snake fauna. For this reason, we recommend intensifying the sampling of medically important snakes in the short and medium term, with a focus on regions such as the southern Andes, the Pacific, the Caribbean (especially in the upper Guajira), Orinoquia, and Amazonia.
Nevertheless, ten years after the first assessment by Lynch et al.
[1] that estimated the geographical representativeness of medically important snake
species in the country (Figure 5A-C), researchers and institutions advocating for snake research and conservation have achieved significant advancements in the knowledge of the distributions of Colombian venomous species (Figure 5E-G). On average, they have added 91 new geographical records per department, provided the first official records for 19 of the 32 departments in Colombia, and achieved significant coverage of 87% of the national territory with at least one geographical distributional record (Figure 5D-H). For accurate and complementary information, please consult:
https://ofidismo.ins.gov.co.
.png)
Figure 4. Sampling effort per pixel of 1.0 geographic degree (110 x 110 km; WGS84) in Colombia during 2010-2024. (A) Representativeness of viperids and elapids species. (B) Representativeness of viperids. (C) Representativeness of elapids. (D) Proportion of viperids/elapids+viperids.
.png)
Figure 5. Comparisons between records reported in this book and Lynch et al,
[1]. (A-C) Sampling effort per pixel of 1.0 geographic degree (110 x 110 km; WGS84) reported by Lynch et al.
[1] for venomous snake species, viperid and elapid species. (D) Proportion of viperids/elapids+viperids per pixel according to Lynch et al.
[1]. (E-G) Advancement in knowledge per pixel for venomous snake species, viperid and elapid species, respectively. (H) Comparisons per political unit between records reported in this book and those by Lynch et al.
[1], showing the advancement in the knowledge of the geographic distribution of Colombian venomous snakes.
2.3 Venomous snake species out of the list
Dubious and puzzling species
Bothriechis schelegelii.— Arteaga eta al.
[16] recently reviewed the
B. schlegelii species complex (i.e.
B. schlegelii and
B. supraciliaris), splitting it in 10 species, five of which were therein described in the publication. Despite our acknowledgement that
B. schlegelii is a species complex, herein we refrain from using their taxonomy due to multiple inconsistencies in the species delimitation and diagnosis. One major problem is that none of the lineages they recognized as species have strong support in their phylogenetic tree (posterior probability values: Colombian node <75%-94%; Colombian intranode <50% pp). Also, the genetic distances between some of the closely related lineages proposed by them as species have genetic distances below the 4% boundary that they proposed as species delimitation criterion.
In their publication, morphological boundaries between closely related and distributed lineages are unclear, morphological characters are ambiguous and feeble when distinguishing their defined species from
B. schlegeliisensu stricto (i.e., see Table 2 in Arteaga et al.
[16] including diagnostic characters as follow: Interoculolabials, canthal scale condition, gular scale condition, loreal in contact with preocular). Although Arteaga et al.
[16] (see supplementary material 1 in Arteaga et al.
[16]) examined 45 characters and 400 specimens, almost 40% of their morphological matrix corresponds to missing data (meaning their actual sample size is ~160 specimens; so, a considerable portion of these characters was not examined for all specimens). Morphometric and meristic characters were poorly described, since only ranges were presented, without specifying the measures of central tendency and the presence of outliers. Likewise, most of these characters require a morphometric analysis to properly state the size and relative proportions among head scales and their diagnostic capacity. Besides, the use of diagnostic characters that depend on sample size (e.g., loreal in contact with preocular) provokes significant biases in species delimitation, as well as doubts about their geographic distributions (e.g., between
B. khwargi and
B. klebbai).
Color pattern is not suitable as a diagnostic character in the
B. schlegelii species complex, as it is widely known to have several biases and constraints, such as ontogenetic shifts
[26], and polychromatic intrapopulation variability
[25,26,39]. Indeed, Arteaga et al.
[16] are aware of this because most of their figures and descriptions of Colombian
B. schlegelii populations clearly showed both ontogenetic and polychromatic variability. Despite this, they insisted on the use of these color patterns as diagnostic characters, even though their diagnostic capacity is low or null.
In Colombia, Arteaga et al.
[16] proposed six species in their work:
B. khwargi,
B. klebbai,
B. rahimi,
B. rasikusumorum,
B. schlegelii, and
B. torvus. Recently, Reyes-Velasco
[149], critiqued these taxonomic changes indicating that the genetic and morphological data fail to support the distinction of these species. Reyes-Velasco argues that the over-splitting of species due to deep intraspecific genetic variation has led to broader taxonomic inflation, as this variation may reflect clinal differences rather than species boundaries.
Reyes-Velasco
[149] concludes that the new species proposed by Arteaga et al.
[16] within the
B. schlegelii group may represent geographic variants or subspecies rather than distinct species. As shown above, Reyes-Velasco’s assessment supports our findings, affirming our decision to recognize only three valid species within the
B. schlegelii complex:
B. nigroadspersus,
B. schlegelii, and
B. supraciliaris. The remaining taxa proposed in recent revisions, including
B. khwargi,
B. klebbai,
B. rahimi,
B. rasikusumorum, and
B. torvus, do not withstand more rigorous analysis and should be synonymized with
B. schlegelii. This result has important implications for conservation efforts in Colombia and underscores the need for caution when making taxonomic changes based solely on mitochondrial DNA. Additionally, unstable taxonomic changes in a medically significant species like
B. schlegelii, due to its association with snakebites (see Chapters
5 and
9), may complicate non-taxonomic literature, including medical and biochemical references.
We recommend that future publications dealing with the
B. schlegelii complex refer to it as we suggest (B. schlegelii), and to be rigorously described the geographic origin of the reported specimens (e.g., accurate locality, coordinates). In this way, when lineages within this species complex are properly delimited, the already published data can be linked to the corresponding lineages or species.
Bothrops ayerbei and
B. rombeatus.— Folleco-Fernández
[76] tried to clarify the
Bothrops asper species complex from the western slopes of the Cordillera Occidental by proposing
Bothrops ayerbei and
B. rombeatus as new members of this complex. However, the description lacked the detailed characteristics needed to clarify boundaries between these proposed species, as well as between the specimens of the type locality (Obispo, Darién, Panama). The descriptions of the specimens´ physical traits were vague, morphological analysis was misconducted, and there was no phylogenetic analysis performed. Hence, taxonomic decisions are difficult to delimit either species.
Also, there was no designation of a type specimen or a formal description of the new taxon by García-Piedrahita
[17]. Ramirez and Solari
[77] point out that
B. rhombeatus is a
nomen dubium because of the lack of type material, also its troubled history that hampers a correct taxonomic assignation due the lack of crucial data on geographical distributions that hinders an understanding of possible interbreeding between
Bothrops ayerbei and
B. rombeatus. Additionally, according to Ramirez and Solari
[77],
Bothrops ayerbei constitutes an unavailable name following to the rules of the current International Code of Zoological Nomenclature, because the code does not accept the publication of new taxa in electronic journals before 2011.
The significance of distinguishing between these taxa as valid species was not properly discussed by Folleco-Fernández
[76]. Despite this, some researchers have used molecular markers to study the venomic variation and antivenomic responses within the
B. asper species complex, using genetic sequences of populations allocated as
B. ayerbei and
B. rhombeatus without a proper taxonomic delimitation or review of these specimens
[36,78,79]. Besides, these authors have not provided accessibility to the official gene databases for these sequences (e.g., GenBank, EMBL). In fact, the only research that analyzes the relationships among the three species found that
B. asper is paraphyletic in relation to the other two
[80]. Therefore, we suggest that future research includes a broad and exhaustive sampling methodology, considering a larger number of individuals from diverse habitats and employing an integrative taxonomic approach that combines more lines of evidence, allowing for a clear delimitation of these species.
Micrurus hemprichii species complex.— Historically,
Micrurus hemprichii,similarly to most of the South American coralsnakes, has suffered several nomenclatural changes since its establishment. Bernarde et al.
[81] provide a comprehensive historical summary of the nomenclatural changes for
M. hemprichii,. Since Peters and Orejas-Miranda
[15] there are two recognized valid subspecies,
M. h.hemprichii and
M. h. ortoni Schmidt
[82], bothdistributed in Colombia. Micrurus h.hemprichii ranges from the Colombo-Venezuelan savannas of the Orinoquia region to rainforests of Guiana, while
M. h. ortoni is distributed across the Amazonian slopes of Colombia, Ecuador, Peru, and Pará in, Brazil. Feitosa et al.
[83] in a conference presentation that was not peer reviewed, proposed to elevate both subspecies as full species taxa. Valencia et al.
[84] followed this proposal, and based on the morphological affinity states that Ecuadorian populations can be allocated to the
M. ortoni definition without providing a taxonomic assessment.
Currently, there is not available a comprehensive taxonomic assessment available that clearly distinguishes the subspecies of
Micrurus hemprichii or populations across its whole known geographic distribution. In addition, previous proposals have not included Colombian populations, despite the fact that the type specimen of
M. h.hemprichii came from an unknown locality of the Orinoquia region; nor have specimens from the type locality of
M. h. ortoni from the eastern versant of the Andes in Pebas, Loreto, Perú been included. Therefore, we recommend that future publications dealing with the
Micrurus hemprichii complex refer to it as we suggest (Micrurus hemprichii), waiting for a rigorous study that helps to elucidate this species complex.
Despite this, Ayerbe-Gonzáles et al.
[85], without proper taxonomic delimitation or review of the specimens involved, reported the first snakebite envenomation caused by
M. ortoni (=Micrurus hemprichii) in Colombia. The cases occurred in two localities of the cis-Andean region of the country. The first case was reported from the eastern slopes of the Cordillera Oriental in the urban area of the municipality of Pajarito, Boyacá. The second case occurred in the urban area of the municipality of Cartagena del Chairá, Caquetá, a settlement located in the floodplain of the Caguán River. Both patients were women who were bitten on the feet, and they presented symptoms such as intense pain in the affected limb that radiated to the lumbar section in the first case. In the second case, the pain radiated to the knee during the first hour of envenoming, reaching the lumbar section after three hours. After 24 hours of envenoming, the patient in the first case exhibited clear neurotoxic symptoms such as mild Rosenfeld’s facies, palpebral ptosis (drooping upper eyelid), bradylalia (slow speech), and continued intense pain presenting hyperalgesia (increased sensitivity to pain) and hyperesthesia (increased sensitivity to sensory stimuli). In contrast, the patient in the second case lacked neurotoxic symptoms and was discharged after a few hours of observation; she was prescribed acetaminophen and cephalexin.
Leptodeira annulata species complex.— The species of the genus
Leptodeira are widespread in American. This genus is composed by 16 nominal species that primarily inhabit lowland dry forests, pre-montane forests and gallery forests in most the natural and disrupted habitats of the tropical lowlands, distributed from southern North America to northern Argentina
[29,86–88]. The close resemblance in color pattern and antipredator displays (e.g., body like S‑coil posture, head posterolaterally expanded simulating a triangular shape) of
Leptodeira species causes these snakes to be commonly mistaken for
Bothrops species in tropical regions. All species are nocturnal, with semi-arboreal habits occurring near slow-moving streams or standing water. These snakes have generalist feeding habits, mainly consume mainly small frogs and lizards, frequently anuran eggs and tadpoles, snakes, and occasionally consume small birds and fishes
[89–92].
Across the whole distribution of the
Leptodeira species, the taxonomic status of species populations has historically been incessantly controversial, and Colombian populations are not the exception. According to Duellman
[86] there are two species in Colombia of
Leptodeira:
Leptodeira annulata with three subspecies (L a. annulata,
L a. ashmeadi,
L a. ashmeadi+rhombiera) and the species
L. septentrionalis with a single subspecies
L s. ornata. The diagnostic characters allowing identification of the two
Leptodeira species are found in its hemipenial morphology, and differences between subspecies are based on color pattern and some scale counts (e.g., dorsal scale reductions). Duellman
[86] also indicates that these species exhibit allopatric distributions in which
Leptodeira annulata populations inhabit the lowlands of cis-Andean ecosystems, while
L. septentrionalis is found in the lowlands of trans-Andean ecosystems in Colombia.
Daza et al.
[93] tested the hypotheses stated by Duellman
[86] using molecular evidence and found paraphyly between
L. annulata and
L. septentrionalis, so that genetic distance did not work as a criterion for species delimitation. However, these authors did not provide a comprehensive evaluation of Colombian populations of
Leptodeira because they focused their sampling on only a few Andean localities (Antioquia, Caldas, Meta [foothills]). Subsequently, Barrio-Amorós
[94] provided a comprehensive taxonomic reassessment and taxonomic acts. This author states that
L. annulata,
L. ornata,
L. ashmeadi are full species distributed in Colombia. However, his proposal lacks care, precision, and sufficient information about the methods and datasets used, creating taxonomic instability due to unfounded taxonomic decisions and untidy descriptions. Therefore, most of his conclusions and decisions should be carefully revised and used conservatively.
Torres-Carvajal et al.
[88] brought taxonomic stability after Barrio-Amorós
[94], by performing a comprehensive and well-supported revision of
Leptodeira snake populations of western Ecuador. They combined molecular and morphological evidence, including the molecular Colombian samples previously utilized by Daza et al.
[93], but they did not included morphological or hemipenial assessments of Colombian populations. These authors presented a maximum likelihood tree that retrieved with strong support the
L. annulata/septentrionalis complex reported by Daza et al.
[93], including Colombian populations.
In addition, within deeper nodes of their tree topology, they retrieved some clades considered as moderately to strongly statistically supported. Among these lineages Torres-Carvajal et al.
[88] made the following taxonomic decisions: (1) based on molecular and morphological evidence they described a new species (Leptodeira misinawui) from a population previously considered as
Leptodeira s. larcorum
[86], and located at moderate to high elevations (950–2,734 m asl) in the southwestern Ecuadorian Andes; (2) based on molecular and morphological evidence solely from Ecuadorian populations, they elevated to full species status the populations previously considered as
Leptodeira s. ornata from western Ecuador and Colombia, central and eastern Panama, as well as the Darien Mountain slope;(3) based solely on molecular evidence from the same populations previously considered by Daza et al.
[93] they elevated to full species
Leptodeira a. ashmeadii.
Costa et al.
[95] reassessing the systematics of
Leptodeira expanding upon and improved the molecular and morphological sampling and provided a new proposal for the polyphyletic
Leptodeira species complex for several South American regions. These authors proposed four species distributed in Colombia:
Leptodeira annulata,L. approximans, L. ashmeadii, and
L. ornata, and they partially agreed with the taxonomic acts of Torres-Carvajal et al.
[88] that recognize
Leptodeira larcorum and
Leptodeira ornata as full species, the latter distributed from western Ecuador, through the Pacific Colombian ecoregion to eastern Panama.
Nevertheless, Costa et al.
[95] indicate that according to their tree topology results
L. ornata is composed of three different taxonomic units (L. ornata 1distributed in distributed in Colombia, southern Panama [type locality of
L. ornata is Isthmus of Darién];
L. ornata 2 distributed in Costa Rica and northern Panama;
L. ornata 3 distributed in Ecuador and Peru).
Costa et al.
[95] conclude that the populations morphologically described by Duellman
[86] from the Darien region in Panama (L. ornata type locality included) and from the Magdalena and Cauca valleys and Chocó region in Colombia are combined for redefined
L. ornata. However, they did not contrast molecular or morphological evidence between specimens from the interandean valleys of Colombia and the Chocó region; therefore, they restricted the distribution of
L. ornata to the Pacific ecoregion of Colombia and southern Panama. However, Costa et al.
[95] in their redefinition of
L. ornata (see the appendix S2 in Costa et al.
[95]) depict the older distribution proposed by Duellman
[86] for
L. s. ornata encompassing most of the trans-Andean region despite the several diagnostic characters employed to distinguish between
L. ornata and
L. ashmeadii that are strongly overlapped between these populations (e.g., color pattern of the head dorsal surface, dorsal scale counts, supralabial scale count). Thus, morphological distinctiveness of these two populations remains uncertain and awaits further studies.
Similarly, Costa et al.
[95] proposed a taxonomic arrangement for the
Leptodeira annulata ashmeadii/bakeri group of species, supporting the previous results of Daza et al.
[93]. The redefinition of Costa et al.
[95] of
L.ashmeadii locate this species in both the Caribbean coast and cis-Andean region of Colombia. However, morphological distinctiveness within Colombia population remains uncertain requiring further studies. Particularly, because hemipenial morphology of the species in the genus
Leptodeira proposed by Costa et al.
[95] both from trans and cis-Andean have no significant differences between each other (Figure 4; see also the appendix S2 in Costa et al.
[95]), and several diagnostic characters employed by these authors to distinguished between them are strongly overlapped.
An ongoing comprehensive study of the hemipenial morphology of the genus
Leptodeira, with special interest in Colombian populations (Angarita-Sierra, unpublished data), shows that the hemipenial architecture of
Leptodeira ashmeadii exhibits conspicuous morphological differences between populations that do not match the proposal of Costa et al.
[95] (Figure 6 A-D versus H-I), while others apparently do fit what is described by these authors (Figure 6 A-B versus C-D). Similarly, hemipenial morphology of trans-Andean Colombian populations (including western Colombia) mismatches the hemipenial architecture of
L. septentrionalis (sensu stricto) described by Duellman
[86], as well as the hemipenial morphology described in
Leptodeira ornata by Torres-Carvajal et al.
[88]. Moreover, the hemipenial architecture of trans-Andean Colombian populations of
Leptodeira ornata proposed by Costa et al.
[95] showed marked variability between sister lineages, as well as within populations of this lineage distributed in Colombia (Figure 6E-G versus K-M).
.png)
Figure 6. Hemipenial morphology of some populations of
Leptodeira annulata species complex from cis and trans-Andean Colombia regions. Sensu Costa et al.
[95]. (A-B):
L.ashmeadii (TAS 689) from Finca El porvenir, Vereda La Colombina, Paz de Ariporo, Casanare; (C-D):
L.ashmeadii (JDL 30721) from Finca El Vogal, Vereda Las Flores de Jaime Botero, El Retén, Magdalena; (E-G)
L. ornata (JDL 29306) from Tumaco, Nariño; (H-J)
L.ashmeadii (JDL 30754) from hacienda La Maria, El Retén, Magdalena. (K-M):
L. ornata (JDL 29872) from Vereda Santa Paola, San Martin, Cesar.
So, despite enormous efforts to disentangle the taxonomy and relationships of
Leptodeira, as well as the significant advancements made by Costa et al.
[95] (including the strongly supported lineages found by them), several problems remain that continue to cause controversy on a fine geographic scale when delimiting the distributional ranges of the genus
Leptodeira. Future studies require broader molecular sampling and a detailed revision of hemipenial morphology within Colombian populations. Considering the difficulties in determining distribution limits and distinguishing species by external morphology, as well as the limited knowledge about venom variation and snakebites caused by these snakes, we have decided to treat all members of the genus
Leptodeira in Colombia as a single species,
Leptodeira annulata, for the purposes of this publication. We hope that future evidence will help clarify the status of Colombian populations of this genus.
Leptophis ahaetulla species complex.— The neotropical genus
Leptophis encompasses a total of 19 snake species distributed from Central America to northeastern Uruguay and Argentina. Four species inhabit Colombia
[29]. Species from this genus are generally diurnal and arboreal, commonly seen in small shrubs and trees, and characterized by their colorful appearance
[96]. They feed mainly on hylid frogs, lizards, and young birds
[97]. When these snakes are threatened, they display a histrionic and aggressive behavior opening the mouth as much as they can, and following the movements of the object that threatens it
[98].
Currently, between the nine
Leptophis species recognized for Colombia three are not taxonomic conflicts, they are well-delimited and are supported by morphological and molecular evidence allowing distinctions between each other (L. cupreus,
L.
depressirostris,
L. riveti). Nevertheless, within the
Leptophis ahaetulla species complex significant differences have been detected across its wide distribution from Central America to northeastern Uruguay. Currently the
Leptophis ahaetulla species complex includes 11 species, six of which were formally recognized in Colombian territory (L. ahaetulla sensu stricto, L. bocourti,, L. coeruleodorsus, L. nigromarginatus, L. occidentalis, and
L. urosticus)
[15,42,101,102,150].
Recently, two major efforts were made to understand the phylogenetic relationships and to put in the taxonomy of the
L. ahaetulla species complex. The first is the most comprehensive phylogenetic study to date
[102], where six of the 12 species of the complex were included. Nevertheless, it should be considered that the hypotheses of the relationships within
L. ahaetulla species group by Torres-Carvajal and Terán
[101] was assessed with a limited taxonomic and geographic survey (this study did not incorporate any Colombian specimens) and have many poorly supported nodes (including the node that encompasses the entire species group), so it is likely that the relationships between species will change when more species or genomic regions are incorporated in future analyses. Even so, this study yields interesting results such as the polyphyly of at least three species (L. ahaetulla,
L.
nigromarginatus and
L. occidentalis) and the data suggests that research be conducted soon to resolve the phylogenetic relationships and the taxonomic status of the group.
The second important initiative for consolidating the current taxonomy of the
L. ahaetulla species group was the revision based on external and hemipenial morphology by Albuquerque and Fernandes
[150]. This research establishes the six currently recognized species of the
L. ahaetulla species complex in Colombia. The research analyzed 71 Colombian specimens of four species (L. ahaetulla,
L. nigromarginatus,
L. occidentalis, and
L. urostictus). Likewise, the recognition of
L. bocourti in Colombian territory is based on the synonymization of
Leptophis occidentalis insularis from Gorgona Island (Cauca department) with
L. bocourti by Oliver
[102], although the type specimens of
L. o insularis were not examined by Albuquerque and Fernandes
[150]. The recognition of
L. coeruleodorsus is based on an anecdotal specimen of
L. coeruleodorsus collected by William W. Lamar and illustrated in Campbell and Lamar
[26] in Villavicencio, Meta
[101].
Although this article was crucial in establishing the current nomenclature of the
L. ahaetulla species complex, there are still many issues such as the limits of species distribution at the national scale that need to be resolved. For example, there is no information on the distributional gap of
L. occidentalis from the tropical dry forest of Guayaquil (Ecuador) from the northern Chocoan Region of Colombia or comments as to whether continental populations of
L. bocourti exist. Considering such biogeographical issues and the epidemiological interests of this work, we prefer to treat all species of the complex as
L. ahaetulla.
Oxybelis aeneus and
O. fulgidus.— The genus
Oxybelis is composed of ten rear-fanged species native to the subtropical and tropical regions of the Americas often associated with defensive and aggressive behavior. Currently in Colombia have been recognized three species
[29]. Herein, we only focused in
Oxybelis aeneus and
O. fulgidus species groups that are considered medically important because they have snakebites reports (i.e., in Brazilian socioecosystems) or have the potential to cause snakebites due their behavior (see Chapter 4).
Oxybelis aeneus is a strictly diurnal snake with arboreal habits and an ambushing foraging strategy
[103,104]. It feeds mainly on lizards (Anolis sp.) and occasionally frogs. However, birds, small mammals, insects, and fish have also been reported as part of its diet
[104,105]. Similarly,
Oxybelis fulgidus is strictly diurnal, spending most of the time on trees or shrubs
[106]. This species is more associated with forest environments and sometimes can be found foraging on the ground
[106]. Its diet appears to be more varied compared to
O. aeneus, consisting mainly of lizards and birds at similar rates
[107–111].
Historically,
Oxybelis aeneus has been considered as one species across its wide distribution range from southern Arizona (USA) throughout the Centro and South America into southeastern Brazil. However, recent studies have sorted out the species crypticity of the brown vine snake
[112,113]. According to the molecular phylogenetic analysis performed by Jadin et al.
[113,114] within the
O. aeneus complex, the species were found to represent four clearly differentiated lineages across North and Central America. Moreover, northern South American populations of
O. aeneus could represent more than one lineage, as well as several undescribed species. For example,
Oxybelis vittatus has been recently resurrected, and its distribution across the Chocoan region of Colombia is expected
[29,113]. Nevertheless, Colombian populations have not been included in the molecular analyses and they lack comprehensive morphological studies. Thus, these populations remain unclear. Due to a lack of new evidence that might help to understand the
O. aeneus species complex inhabiting Colombia, we retained all the Colombian populations under the name
Oxybelis aeneus.
Colombian populations of
Oxybelis fulgidus exhibit similar taxonomic issues as the
O. aeneus species complex
[112], as well as the lack of new lines of evidence that allow us to perform species delimitation analyses to sort its species crypticity. Therefore, we retained all the Colombian populations under the name
Oxybelis fulgidus.
Wrong or troubled localities, and dubious species determination
Due to the complex topography of Colombia and the lack of accurate knowledge about fine-scale geographic distributions of venomous snakes, several records have been reported with high uncertainty. Over time, these records have been considered 'valid' or probable distributions, without any researcher providing evidence to support their validity. Similarly, edges between ecoregions, in which there might or might not be hybridization zones between widespread species (e.g.,
Bothrops asper and
B. atrox) provide puzzling distributional records because taxonomic determination is dubious.
Previously, Campbell and Lamar
[26] pointed out some of these areas across Colombia, indicating that further studies should be done to elucidate the suitable distribution of Colombian venomous species. Our review detected several problematic localities, as well as dubious species determination of several venomous species in the Colombian area that match the controversial regions indicated by Campbell and Lamar
[26], but we were also be able to identify others records and regions of controversy (see
https://ofidismo.ins.gov.co). Since the Campbell and Lamar
[26] distributional remarks few studies have addressed these concerns
[35]. We suggest that, due to the lack of a proper revision of these records, they should be carefully used in future studies that seek to model species distributions, species niches, and ecological traits employed as characters in species diagnosis and delimitation, as well as models that estimated the envenoming snakebite risk. Below, we provide a brief description of the main wrong or troubled localities of several venomous species in Colombian.
Bothrops asper-atrox complex.— Campbell and Lamar
[26] indicate that confusion surrounding
Bothrops asper and
B. atrox range distributions arose since Garman in 1883
[115] recognized both
B. asper and
B. atrox but confused these species both with each other and with other congeners. In Colombia, the area of controversy encompasses the foothills and uplands of the central and northern eastern slopes of the Cordillera Oriental in the municipalities of Choachí, Fómeque, Ubalá, Quetame, Guayabetal, San Antonio del Tequendama, Tena (Cundinamarca), Paya, Pajarito, San Luis de Garagoa (Boyacá), Catatumbo, Pamplona, Sardinata, and Toledo (Norte de Santander). Similarly, there are several problematical localities relating to their elevational ranges. Both
B. asper and
B. atrox,throughout their known geographic distributional ranges, occupy the lowlands (<1000 m above sea level [hereafter asl]).
Nevertheless, both
B. asper and
B. atrox have records that exceed their known elevational maximum range (Table 1). Problematical records of
Bothrops asper over 2,200 m asl can be found in municipalities such as Santa Rosa de Osos, San Roque, Yarumal (Antioquia), San Lorenzo (Magdalena), and Calima (Valle del Cauca). Dubious
Bothrops atrox elevational records have been reported in Quetame (Cundinamarca) and Pamplona (Norte de Santander). These records could result from misidentification, due to confusion with similar congeners (e.g.,
Bothrocophias species), or undescribed species. Recently, studies have provided some evidence that both
B. asper and
B. atrox are polyphyletic and could hide unnamed lineages (see Chapter 3) across Colombian ecoregions, but additional efforts must be made to untangle these taxonomic problems and allow the clarification of their accurate range distributions.
Bothrocophias colombianus.— The enigmatic toadheaded pitvipers of the genus
Bothrocophias are some of the medically important, yet poorly known, South American viperids. These snakes inhabit isolated and difficult-to-access ecosystems in South America and are poorly represented in biological collections
[116]. In particular, the known geographic distribution of
B. colombianus is restricted to the hyperhumid Chocoan rainforest in the departments of Chocó, Cauca, and Risaralda. However, there is a problematic locality in Yarumal, Antioquia department. This could likely be a case of species misidentification because it is too far from the historically known geographic distribution. The specimen that supports this record was not reviewed; it is housed in the reptile collection of the Museo de La Salle (Bogotá) under catalog number MLS 1832. The geographic record was retrieved from the Sistema de «Información Biológica Colombiano» (SiB Colombia, Spanish acronym).
Crotalus durissus.— This South American rattlesnake, across its known geographic distribution range, mainly occupies lowlands <1,000 m asl (Table 1), but there are some Colombian localities on the western slope of the Sierra Nevada de Santa Marta (Magdalena) that are remarkable elevational confirmed records at 1,700 m asl, exist in localities such as San Lorenzo. However, there are two records from upland that are outside from its historically known range distribution. One is from the vereda El Centro, Villa de Leyva (Boyacá, IAvH 4849), and the second is from Fusagasugá at 1,729 m asl (Cundinamarca, CRODUR00021 INSZ collection). However, both records likely resulted from human activities causing a translocation from the original range distribution area to the reported locality.
Hydrophis platurus.— The Yellowbellied Sea snake is the only sea snake present in Colombia, and probably in the entire Western Hemisphere. Hernández-Camacho et al
[117] states thatthere areColombian populations of
H. platurus present in four localities of the Caribbean coast [Ciénaga de la Virgen (Bolívar); between Punta San Bernardo and San Bernardo archipelago (Sucre); SE Fuerte island (Córdoba); and near to mouth of the Canal del Dique in the Bay of Cartagena (Bolívar)]. However, these authors did not provide collection numbers for the specimens, collector numbers or vouchers that support their findings, neither did they provide pictures or any documentary evidence of the new records. Indeed, they did not provide any information about the origin of the records or how they got them. In contrast, these authors provide a comprehensive speculative analysis of three possible explanatory hypotheses about the expansion of distribution expansion of
H. platurus into the Caribbean Sea, but this analysis is not supported by evidence that directly links the distributional expansion of
H. platurus.
Currently, the controversy about the distributional expansion of
H. platurus into the Caribbean Sea continues because informal communications from fishermen and marine biologists indicate that
H. platurus could be present in some localities of the Colombian Caribbean Cost, but no evidence has been provided yet. Thus, a comprehensive revision of the specimens housed in biological collections and fieldwork sampling must be conducted to determine whether the distribution for this species has reached the Caribbean Sea.
The most likely route of translocation is through ships’ ballast water. Ballast water is one of the major pathways for the introduction of nonindigenous marine species
[117,118]. Due to the continuous shipping between the Caribbean Sea and the Pacific Ocean across the Panama Canal, it is possible that
H. platurus might have expanded its distributional range into some localities of the Colombian Caribbean Coast using ballast water, and this is a possible explanation for the snakebite cases informally reported in this region. Nevertheless, further studies must be developed to obtain reliable information about the epidemiological pattern of this snakebite event, as well as to assess the ships’ ballast water hypothesis (see Chapter 9).
Micrurus dumerilii.— This coralsnake is distributed in the South American mainland and restricted trans-Andean ecoregions of Colombia. However, there are two records outside its known historical range distribution. One is from the San Andrés Island (Archipelago of San Andrés, Providencia, and Santa Catalina) located about 230 kilometers east of mainland Central America and 750 kilometers north of mainland Colombia. The second is from El Encanto (Amazonas). However, both records are likely to result from dubious species determinations, due to confusion with similar congeners (e.g.,
Micrurus tikuna for the Amazonian record).
Micrurus isozonus.— This is a widely distributed in the cis-Andean lowlands of northern South America in the Guiana Shield and Orinoco regions and the northernmost portion of the Amazon Basins
[119] Recently, a controversial record of
M. isozonus was documented from the Caribbean region of Colombia. Tavares-Feitosa et al.
[119] reported one specimen housed in the Natural History Museum of Paris, which was collected in Cartagena, Bolívar, Caribbean lowlands. This unexpected record is located approximately 700 km in a straight line from the nearest record, in northwestern Venezuela. Nevertheless, there are no more records of this species along in the Colombian Caribbean, which is one of the best inventoried regions in the country. When reviewing the museum online catalog, the available information states that the specimen is of unknown provenance. Considering that there is no evidence that this specimen comes from Cartagena, this record will not be included in this book.
Micrurus multifasciatus.— This bicolored coralsnake occurs in lower Central America from Nicaragua to Panama, inhabiting lowland moist and wet forests (rainforests), subtropical wet forests, and lower montane wet forests
[26]. Castro-Herrera and Vargas-Salinas
[120] reported this coralsnake from western Colombia in Valle de Darien, Rio Azul, 7 km from the camp “Campo Alegre” (UVC 6676), extending its geographic distributional range by approximately 419 km (airline) from its known southernmost locality in Panama (Central America). After reviewing this specimen, we noticed that it corresponded to
M. multiscutatus a snake species endemic to Colombia and restricted to the Pacific region and that had been confused with
M. multifasciatus (Figure 7), a remarkably similar species. Therefore, although
M. multifasciatus is expected to be distributed in Colombia in the Chocoan ecoregion, there are currently no confirmed records of it.

Figure 7.
M. multiscutatus. (A-B). Specimen the
M. multiscutatus. from western Colombia in Valle de Darien, Rio Azul, 7 km from the camp “Campo Alegre” (UVC 6676), pictures by Carlos Andrés Linares. (C)
M. multiscutatus in life, and (D-E) fresh euthanized from type locality: El Tambo, Cauca, Colombia. Pictures by Luis Vera-Paez.
Micrurus nigrocinctus.— This widely distributed species occurs from southwestern Mexico to northwestern Colombia, for which a deeply cryptic diversity has already been demonstrated
[121]. A controversial record of
M. nigrocinctus from Old Providence, Colombian Caribbean islands, was based on one specimen collected by CH Towsend in 1884, but it was lost in 1921 in a shipment from the United States National Museum (Washington, USA) to the Museum of Comparative Zoology (Cambridge, USA)
[122–124]. The snake has not been recorded in Old Providence since the late 19th century, so there is some doubt as to whether the locality of the lost specimen was correct. At worst, if the snake did exist in Providence, it is now extinct.
Micrurus putumayensis.— Currently, there are some snake species lists that include
Micrurus putumayensis among Colombian coralsnakes (i.e., Reptile database
[29]), but it appears that this was due to a "broken phone" effect in the interpretation of information over time. Campbell and Lamar
[26] report for the first time
M. putumayensis in Colombia based on the list of Colombian snakes reported by Medem
[21]. However, Medem
[21] indicates there are not records of
M. putumayensis in Colombia, but he suggests that likely some specimens and localities of snakes alike
M. steindachneri (=
Elaps fassli Werner 1927) could be confused with
M. putumayensis by Werner
[125]. The type locality of
M. putumayensis is Puerto Socorro (Peru?) 270 Km. NE from Iquitos. However, after looking for it at digital geographic gazetteers (Geonames, Google earth) and Colombian and Peru official maps, this locality is unknown. Thus, the distribution of
M. putumayensis species does not include Colombia, being restricted to Brazil and Peru.
Porthidium lansbergii-nasutum complex.— In Colombia,
P. nasutum and
P. lansbergii are closely related species, characterized by cryptic morphological traits such as a prognathous face or 'hog-nosed snout' character.
Porthidiumnasutum is distributed across the biogeographic Chocoan ecoregion to the Chocó-Darién ecoregion, while
P. lansbergii as well is distributed in the Isthmian-Pacific dry forests in Panama and the Colombian Caribbean, as well as in the Chocó-Darién region, where it transitions into the Urabá humid forests and the evergreen forests and dry forest in the Magdalena Valley (see Chapter 3).
Porthidium nasutum and
P. lansbergii are sympatric in the Chocó-Darién region; thus, in localities such as Apartadó, Carepa, Urrao, Segovía (Antioquia) misidentification due to confusion of these species with each other is common.
Both
P. nasutum and
P. lansbergii have records that exceed their known elevational maximum range (Table 1). Problematic records of
P. nasutum over 1,500 m asl can be found in municipalities such as Urrao (Antioquia), and for
P. lansbergii in localities like Amalfi, Guatapé (Antioquia), Cimitarra, Betulia (Santander), and Ocaña (Norte de Santander) with an elevational range between 1,674-1,823 m asl. All these records require a detailed revision to confirm if they represent real distributional records of
Porthidium species or are misidentifications.
Historically, the presence of
P. lansbergii in the Orinoquia ecoregion has been based on anecdotal and informal reports, but no evidence has been provided; thus, they have been considered as speculative and spurious records. In this book (see Chapter 3), we confirmed the first record of
P. lansbergii from Orinoquia ecoregion. A female (INSV-SR-00288) from Yopal (Casanare) was found by the Fire department during rescue activities in the foothills of the per urban area of Yopal city on July 24, 2021, and was delivered to the Alive Venomous Animal Collection of the National Institute of Health (INSV). Molecular phylogeny analysis highlighted that this specimen was nested within the Caribbean clade, showing low a genetic distance compared to specimens from the Colombian Caribbean coast (see Chapter 3).
3. Endemicity and poorly known venomous snake species in Colombia
Most of Colombia's venomous snakes are by nature cryptic, rarely seen in the wild and apparently having low-density populations. Thus, their endemism or narrow range distributions could be an artifact of our fragmentary understanding of them or might be their true nature. Nevertheless, despite the huge efforts of several researchers conducting fieldwork in their known cryptic habitats and searching through the cabinets of biological collections for uncatalogued or lost specimens, several Colombian venomous snake species remain poorly documented and are represented by few specimens. This circumstance strongly limits our understanding of the essential aspects of their biology or their interaction with human communities. Also, their crypticity has rendered them enigmatic beings, to the point of becoming legendary among herpetologists, who boast of having seen one of these snakes in their lives.
From the total of 49 venomous snake species in Colombia, 16 venomous snake species have ten or fewer known records or localities in Colombia, representing between 0.2-0.02% of the total records available for Colombian venomous snakes (~5,488 records, see
https://ofidismo.ins.gov.co), making them the most enigmatic venomous snakes of the country (Table 2). We highlight some of the species here.
For example,
Micrurus oligonellatus and
M. renjifoi are known for their type localities and are represented by their type series; indeed, no natural history data is known.Micrurus spurrelli is known from five records including the type specimen, no live pictures of this species are available, and no natural historical essential data is known. Since its erection by Boulenger in 1914 its known distribution is restricted to four localities in the Chocoan rainforest of the Colombia
[25].
Micrurus psyches has a controversial distribution. Campbell and Lamar
[26] reported that this species is absent of Colombia, suggesting that records of
M. psyches from the Orinoquia region are questionable and that these specimens could potentially represent an undescribed bicolored coralsnake species mistaken for
M. psyches. However, some specimens housed in the INS reptile collection match with the diagnostic characters described by Roze
[126]. Nevertheless, due to the small sample size, the evidence they provide is insufficient to test any hypothesis. Thus, for this work, we have decided to treat them as
M. psyches, hoping that future evidence will help clarify the status of Colombian populations.
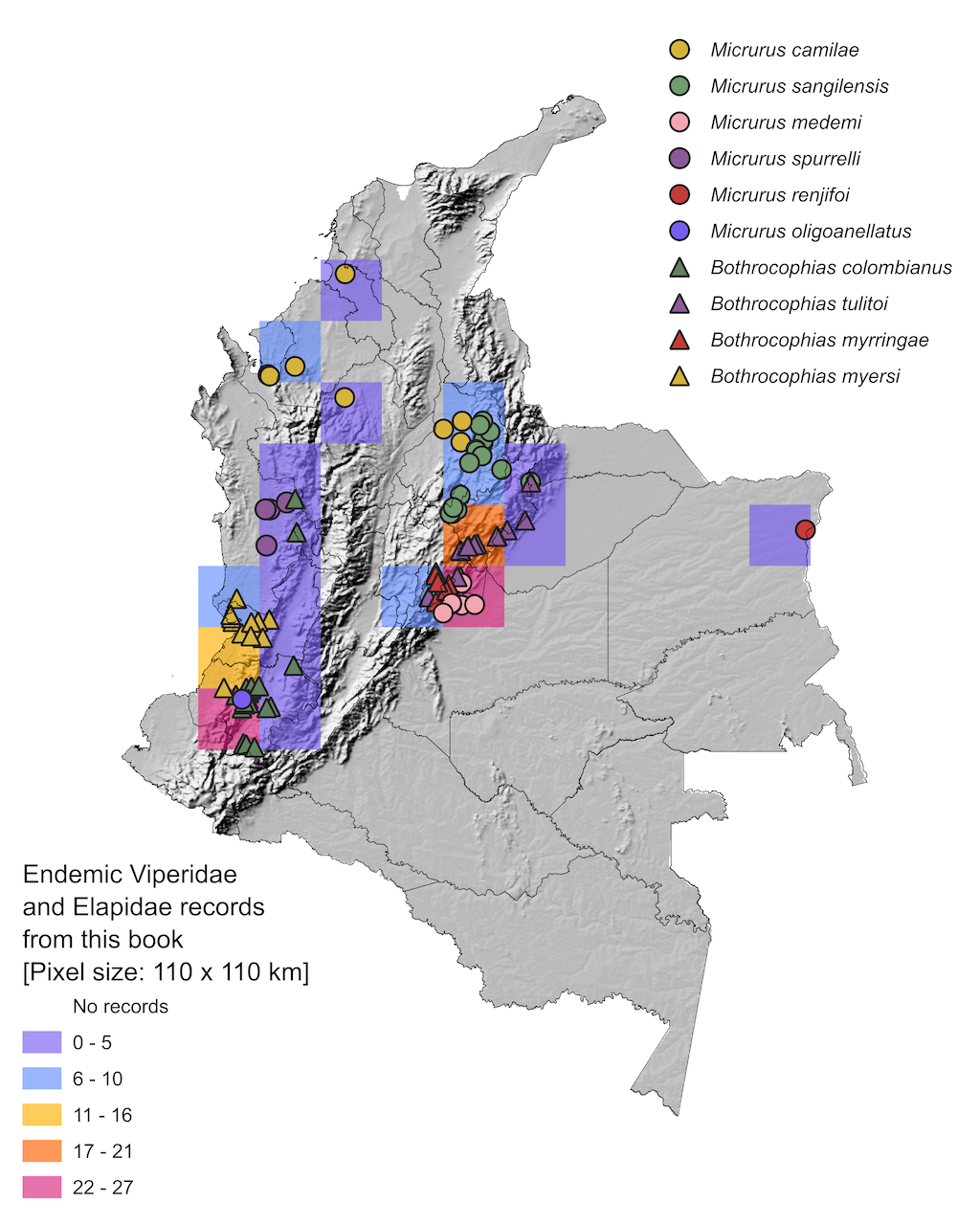
Figure 8. Distribution of endemic venomous snake species in Colombia
Table 2. Poorly known venomous snake species, with unique or restricted records geographically.
| Criteria |
Species |
Geographic distribution |
| Species with singe record or locality |
Micrurus oligonellatus |
Cauca: El Tambo, (Type locality) |
|
Micrurus renjifoi |
Vichada: Puerto Carreño, Tomo River near its juncture with the Río Orinoco (Type locality) |
| Species with between two to five records or localities |
Micrurus nattereri |
Guainia: stream Raya. Vaupés: Yavaraté Monfort; Pamopetá, Canoti. |
|
Bothrops pulcher |
Caquetá: Florencia; San Vicente del Caguán, PNN Cordillera de los Picachos |
|
Micrurus psyches |
Arauca: Arauquita. Meta: Villavicencio, Parte superior del Caño El Buque |
|
Micrurus remotus |
Guainía: Negro River. Meta: La Macarena, Piñalito, Cabaña Paujiles. Vaupés: Mitú. |
|
Micrurus multiscutatus |
Cauca: El Tambo (Type locality). Valle del Cauca: Valle de Darien, Rio Azul, 7 km from the camp “Campo Alegre” |
|
Micrurus spurrelli |
Chocó: Condoto, Peña Lisa (Type locality); El Carmen de Atrato; Quibdó, Pacurita |
|
Micrurus tikuna |
Amazonas: Leticia, road Leticia-Tarapacá km 7 |
| Species with between six to ten records or localities |
Bothrocophias campbelli |
Chocó: Itsmina, Andagoya. Nariño: Barbacoas, Ñambí, Reserva Natural Río Ñambí, Vereda el Barro, corregimiento de Altaquer. Valle del Cauca: Buenaventura, Bajo Calima |
|
Micrurus clarki |
Cauca: Guapi. Chocó: Condoto; Itsmina, Andagoyá, upper San Juan River; Riosucio, PNN Los Katios. Valle del Cauca: Dagua, La Elsa, road Cali-Buenaventura. |
|
Bothrocophias myrringae |
Cundinamarca: Choachí, Palo Alto; Fómeque, Vereda de Coasavistá; El Calvario; La Calera, Vereda Mundo Nuevo (Type locality); Guayabetal. |
|
Micrurus nigrocinctus |
Antioquia: Carepa; Necoclí; Turbo, Currulao River; |
|
Hydrophis platurus |
Cauca: Guapi, PNN Isla de Gorgona. Chocó: Bajo Baudó, Cabo corrientes; Juradó, Cabo Marzo; Nuquí, Morros de Jurubida. Nariño: Tumaco |
|
Micrurus scutiventris |
Amazonas: Leticia, Leticia; La Chorrera |
|
Bothrops taeniatus |
Amazonas: Leticia, PNN Amacayacu, Río Amacayacu;Puerto Rastrojo Mirití-Paraná River. Cundinamarca: Medina. Guainía: Puerto Colombia,Macanal, Garagon. Vaupés: Pacoa; Taraira, Estación biológica Caparú. Vichada: Cumaribo |
Venomous snake species with narrow range distribution restricted to Colombia, also known as endemic species, represent 4.9% (10 species) of the total medically important snake species in the country (Figure 8). Except for
Micrurus medemi,
M. sangilensis,
Bothrocophias myrringae and
B. tulitoi, all of them have in common that they are inhabiting in isolated difficult-to-access habitats, or areas risky for researcher’s due to the Colombian armed conflict. Thus, any data or information, robust or anecdotical, gathered for these species represents an important achievement.
Currently, social media platforms such as Facebook, YouTube, Instagram, Twitter, iNaturalist, blogs, and wikis have contributed to obtaining information and knowledge about these enigmatic venomous snakes in a cost-effective and quick manner, rather than relying on traditional, expensive, and challenging expeditions to unvisited, cryptic, or difficult-to-access habitats
[3]. Citizen-science initiatives based on local community observers can significantly contribute to an understanding of the poorly known venomous snake species (i.e.,
Micrurus ancoralis geographic range expansion)
[3,127,128]. However, biases associated with rainfall, population, and internet penetration must be considered when this type of data is used
[3].
In addition, although the quality of the snake records retrieved from social media, the information might be sufficient to determine geographic locations, although the taxonomical identification of species could have several shortcomings. Identification becomes challenging when key characters are not visible. For instance, social media posts often only show conspicuous external traits rather than detailed or internal diagnostic characters such as scutellation or hemipenial morphology
[3]. Therefore, we recommend being careful and conservative when using social media records based on human observation as a primary proxy for establishing geographical distributions or assessing ecological niches.
4. Conservation status of the medically important snake species in Colombia
As seen above, previous studies on snakes have focused on public health, taxonomic issues, species lists and descriptions. Since 2007, Colombia has seen the first initiative that focused on snakes as a relevant and important conservation objective. This conservation initiative was led by the Ministry of Environment and Sustainable Development of Colombia (MADS or Minambiente, current Spanish acronym), the National Health Institute of Colombia (INS or Instituto Nacional de Salud, Spanish acronym), the Regional Autonomous Corporation of the Center of Antioquia (CORANTIOQUIA, Spanish acronym), and the Natural Sciences Institute of the Universidad Nacional de Colombia (ICN, Spanish acronym). This initiative generated a workshop named the “First national Research symposium on snake biology and conservation,” which convened most of the academics, stakeholders, governmental entities concerned with wildlife and environmental heritage, and neophytes.
In 2008, a first draft (not peer reviewed) of the Colombian snake conservation program was organized as a report that summarize the main findings and conclusions of the this workshop
[129]. This report was the initial input for ongoing research, initiatives, conservation plans, and social-media initiatives towards snake conservation in Colombia. In 2012, Lynch
[6] provided the first general assessment of the potential threats that snake populations face in Colombia, addressing habitat loss, deliberate killing by humans, roadkill, illegal wildlife traffic, and scientific research as possible main threats.
Based on this work, as well as the previous Colombian snake conservation report, and thanks to a research project supported by a cooperation agreement between Minambiente, INS and ICN, Lynch et al
[1] in 2014 presented and updated a peer-reviewed a national program for the conservation of snakes present in Colombia (hereafter PNCS, Spanish acronym). In 2016 the PNCS was adopted by Minambiente as the national public policy guideline for the conservation of snakes in Colombia. This public policy guideline stated that currently in Colombia the three main threats for snake conservation are: habitat loss, deliberate killing by humans, and roadkill.
In 2015, the first snake species conservation assessments were led by the IUCN (International Union for Conservation of Nature) reptile specialist group and the Instituto de Investigación de Recursos Biológicos Alexander von Humboldt (IAvH, Spanish acronym)
[130]. This assessment based on experts’ knowledge and judgment (~20 national reptile experts), and IUCN criteria used to evaluate if a taxon belongs to the Red List of Threatened Species categories
[131], resulted in the categorization of ten Colombian snakes species, three of them venomous species allocated in two threatened categories:
Micrurus medemi (Endangered),
M. sangilensis (Vulnerable), and
Bothrocophias campbelli (Vulnerable). This assessment has been the main input to public policy resolutions that indicate and rule the state of snake species conservation state of biodiversity in Colombia
[132].
For Colombian snakes, besides the experts’ knowledge and judgment, the main criterion employed to determine if a species is allocated or not into an IUCN threatened category was the geographic range
[31], mainly due to the dearth of information about criteria such as population size reduction, small population size and decline, and quantitative analysis about the probability of extinction in the wild. The geographic range criterion considers two main metrics, the extent of occurrence (EOO) and area of occupancy (AOO), which should be linked together with at least two of the three following conditions: (1) severely fragmented or number of locations; (2) continuing decline of area EOO or AOO (observed, estimated, inferred or projected), and extreme fluctuations in EEO, AOO, number of locations or subpopulations, and number of mature individuals
[131].
Therefore,
Micrurus medemi was categorized as Endangered
[133] by having severely fragmented locations, and an EOO <5,000 km² that it is in continuing decline as well as the AOO; while
Micrurus sangilensis and
Bothrocophias campbelli were categorized as Vulnerable by having severely fragmented locations, and EOO <20,000 km² that it is in continuing decline losing its habitat quality.
Following this criterion, the Colombian threatened venomous species list must be extended to include at least five of the 16 poorly known venomous species listed in Table 2, as well as three endemic species depicted in Figure 8. For example, species such as
Micrurus oligoanellatus,
M. renjifoi, and
M. spurrelli match with Critically Endangered metrics by having EOO <100 km² that a species is in continuing decline losing its habitat quality;Bothrocophias colombianus,
B. myrringae, and
Micrurus camilae match the Endangered criteria by having EOO <5,000 km² that the species is continuing loss of its habitat quality. Additionally, species such as
B. tulitoi match Vulnerable by having severely fragmented locations, and EOO <20,000 km² that it is in continuing decline losing its habitat quality. Therefore, they should also be included.
An updated assessment of the Colombian threatened venomous species is urgently needed because some of the broadly distributed Colombian venomous snake species are facing significant threats. This is the case with
Crotalus durissus, which beyond facing main threats such as habitat loss and deliberate snake killing by humans, is subject to strong illegal trafficking within Colombia. This species is utilized across the country in magical/religious procedures, as well as ingredients of traditional medicine to treat cancer, erectile dysfunction, and as a sexual enhancer.
Therefore,
C. durissus populations are constantly under significant pressure due to uncontrolled trafficking, but the species lacks a comprehensive conservation assessment that helps to address actions and strategies to reverse, mitigate, or stop this threat. Currently,
C. durissus is listed in the appendix III of CITES (Convention on International Trade in Endangered Species of Wild Fauna and Flora) providing it legal protection for trading or traffic when specimens or its parts are exported from Colombia. However, inside Colombia, there are no specific laws or normativity that protect their populations beyond the environmental normative that states the requirements for the licenses for commercial use of wildlife (i.e., Law 84 of 1989; Law 611 of 2000; decree 4688 of 2005; decree 1076 of 2015; decree 1272 of 2016; resolution 1263 of 2006; resolution 1909 of 2017).
Ten years after the publication of the PNCS, the country continues far from the goal of achieving the mission, objectives, and scope proposed in this conservation public policy. This shows that the efforts invested were not enough and we should reinforce them. Despite snakes having the largest number of herpetological studies of non-avian reptile diversity in Colombia
[134], there are few studies addressing their main threats
[135–141]. However, the major advance in the PNCS implementation was achieved in strategy I:
Increase in the level of knowledge about the snakes present in Colombia; and strategy IV: Implement and develop biomedicine and bioprospecting activities with Colombian snake venoms. Currently, the country has gained a lot of undertesting about Colombian snakes, both in general and applied biology, which have provided enough merit and motivation to write and edit a book like the one you are reading right now.
Nevertheless, the strategies of the PNCS focused on environmental and conservation education (strategy II), as well as recovery and restoration of the natural snake habitats (strategy III) have been poorly or non-implemented. Similarly, lines of action advocating for legal, administrative and financial strengthening for biological research on snake conservation were never implemented. Thus, the priorities proposed ten years ago remain in force and without being properly implemented, so habitat loss, deliberate snake killing by humans, roadkill, and illegal trafficking continue being the main threats for snake populations in Colombia.
Despite the lack of implementation of the PNCS, the publication of this conservation public policy over the years has helped to increase the number of groups of academics, researchers, zookeepers, and the general public that engage in snake research and conservation. It has also motivated the development of robust initiatives aimed to change the negative perceptions of Colombians towards snakes. For example, a growing Facebook community focused on snake research and conservation advocacy has spontaneously led to a robust network (> 1.5 million members) that contributes important information to the geographic distributions of snakes, valuable natural history observations for several snake species, knowledge about the conditions of snake–human encounters in rural and peri-urban areas, and promotes the education and understanding about the biology of venomous and non-venomous snakes in Colombia
[51].
To tackle the main threats for snake populations in Colombia, we recommend promoting the implementation of the lines of action of the PNCS, as well as initiatives that focus on ethnozoological, social, and psychological studies about human attitudes toward snakes, as well as educational actions focused on snake-human encounters in wild or rural areas. Understanding the drivers that provoke negative experiences during these encounters will help develop strategies that promote snake conservation and reduce snakebite incidence.
4.1 Two sides of the same coin: Conservation and ophidism
Conservation biology and ophidism (as a tropical neglected disease) have strong convergences in how these disciplines address their own paradigms. For example, both disciplines share at least eight factors or goals that are assessed and valued: threats, vulnerability, risk, behavior, attitude, prevention, mitigation, and location (Figure 9). Besides, both disciplines address their guiding questions for research and plan actions in a similar manner. Therefore, we propose that actions based on an interdisciplinary approach that integrate knowledge, values, and techniques used by both disciplines could reduce the main snake conservation problems, as well as the incidence of snakebite. The conjunction of these disciplines will help build explicit strategies that are communicable, assessable, and have strong applicability.
Figure 9. The eight common factors or goals that are assessed and valued by conservation and snakebite research.
This conjunction between conservation biology and the study of snakebite can be nested into two comprehensive views of the health sciences, known as the OneHealth and Ecohealth approaches
[142,143]. These proposals raised a conceptual framework that could catalyze conservation biology and snakebite as an interdisciplinary approach to face snake conservation and snakebite accidents. Onehealth focused primarily on the interface of humans and domestic animals in specific social and ecological contexts. But it can be extended to wild animals that commonly share rural and peri-urban locations, which snakes share. An Ecohealth approach
[143,144] considers the close linkages between ecosystems, society, and health, incorporating multiple types of knowledge from natural and social sciences, and the humanities, with the involvement of stakeholders at many levels within a participatory research frame
[144]. EcoHealth has a broader scope compared to OneHealth that focuses specifically on the human-animal health interface. EcoHealth aims to achieve a comprehensive understanding of health and well-being that encompasses the humanities as well as the natural, social, and health sciences
[145].
Due to the multifactorial causes of both the decline in snake populations and snakebite incidence, holistic perspectives as shown above could allow better understanding and might aid in building robust lines of action. Currently in Colombia, there are no initiative for snake conservation or snakebite prevention that have used Onehealth or Ecohealth approaches, but interest in these subjects is growing. Despite the broad scope of PNCS, which includes an interdisciplinary approach, its low implementation of their action lines has limited achievements and perspectives for addressing snake conservation and snakebite incidents. Therefore, it is expected that in the near future the integration of conservation biology and snakebite could occur under these conceptual frameworks.
5. Future perspectives
Colombia holds a special spot in the world due to its geopolitical location, historical ecological significance as a tropical country, and evolutionary role as a pathway for interchanges of South and North American biological elements, which facilitated the great immigration and emigration of snake lineages. Additionally, the convergence of three of the most diverse terrestrial ecoregions on the planet—the Andes, Chocó, and Amazon—has promoted the evolution of astonishing species richness, with several diversification patterns and lineages observed in a relatively limited area (see Chapter
2 and
3). As a result, Colombia is one of the 25 most important biodiversity hotspots worldwide
[146], and is classified as a megadiverse country.
This fact presents significant challenges for snake research and the reduction of snakebite accidents. In particular, Colombia faces strong constraints related to poverty, food insecurity, limited access to medical assistance in rural areas, armed conflict, poor implementation of environmental policies, and low state and private investment in science and technology research that addresses the unmet needs of the general population.
Therefore, to achieve high sampling completeness of Colombian snake assemblages and gain the understanding needed to address snakebites as a neglected tropical disease, an interdisciplinary approach is required to tackle the eight factors described above that underlie the relationship between humans and snakes (Figure 9).
This interdisciplinary approach should be addressed by transforming our relationship with nature, as well as our production and consumption practices based on collective actions with the participation of all members Colombia society. The guidelines to undertake this task were defined ten years ago in the mission, goals, and scope proposed in this PNCS conservation public policy
[1]. Particularly, to reinforce the environmental and conservation education (strategy II), as well as a recovery and restoration of the snakes' natural habitats (strategy III) a reduction of the threats for snake populations in Colombia is urgently needed, as well as to mitigate and properly attend snakebites accidents in Colombia (see Chapter 9).
Particularly, the research presented in this chapter highlights the urgent need to establish a research grant program to support field surveys and studies across various disciplines, aiming to enhance our knowledge of medically important snake species and to train researchers to address the challenges associated with venomous and non-venomous snake species. Taxonomic studies and species descriptions in a megadiverse country like Colombia play a substantial role in the conservation of our natural heritage and contribute to the growing emphasis on caring for the ecosystems that sustain us. Encouraging these activities will also enable us to explore and expand our understanding of how snakes can contribute to the development of new medicines and markets based on venoms, as well as the ecosystem services provided by snakes (see Chapter
6 and
10).
Appendix: Material and Methods
Revision of the available lines of evidence
To achieve the objective of providing taxonomic treatment and criteria that help assign problematic snake species to a taxonomic entity based on the available evidence, we employed three approaches. First, we followed the PRISMA methodology
[147] to retrieve from Google academicandWeb of sciencepapers about taxonomic or systematic revisions, as well as species descriptions of the venomous snakes distributed in Colombia according to the Reptile Database Web site
[29]. Second, we consulted, as far as possible, the taxa-related references included in the Reptile Database Web site
[29]. Third, in the cases in which we were aware that some researcher was currently working on taxonomic or systematic revisions of snakes of medical importance in Colombia, we enquired about his/her expert-opinion in order to incorporate it into the taxonomic treatment proposed
[148].
Search methods for identification of studies: Electronic searches
We conducted monthly searches from April 2021 to June 2024 in the following electronic repositories and Web engines: MEDLINE, Scielo, Scopus, Google Scholar and Pubmed. The search strategies contained the following keywords and Mesh were combined in different ways using the boolean connectors AND OR: snake venoms,
Bothrops,
Crotalus,
Bothriechis,
Porthidium,
Lachesis,
Micrurus,
Xenodon,
Leptophis,
Erythrolamprus,
Oxybelis,
Helicops,
Bothrocophias,
Thamnodynastes,
Leptodeira,
Philodryas, taxonomy, distribution, Neotropics, morphology, species delimitation, integrative taxonomy.
Searching for other resources
We checked relevant cited studies while reviewing the reports identified by the electronic searches, as well as reference lists from any directly relevant reviews identified. We did not apply language or date restrictions and included studies regardless of the type of publication (e.g., conference abstract, trial registry entry, journal article, book).
Contrasting lines of evidence with previous taxonomic proposal
We identified 2,234 publications with potential for inclusion. Of these, we identified a total of 320 articles that underwent title and abstract screening; of these, 36 studies were duplicates and 40 did not meet the search criteria, thus all were excluded. A total of 244 articles were included.
We took as baseline the taxonomic treatment proposed by Campbell and Lamar
[26] for snakes of medical importance in Colombia. Then, we contrasted it with new evidence (after 2004) retrieved from searches done aforesaid available from taxonomic or systematic revisions, and finally, we made a taxonomic decision.