1. Introduction
During the last four decades, toxins found in the venoms of the families Viperidae and Elapidae (see Chapters
2 and
3) have been thoroughly studied using compositional, functional, and evolutionary approaches because of their paramount medical relevance (see Chapter
8,
9 and
10)
[1–3]. Viperid and elapid venoms have received enormous attention since they possibly might cause serious and life-threatening envenomation when human populations expand into their natural habitat and increased contacts take place between humans and snakes (see Chapter 9)
[1,4–7]. These studies revealed an incredible diversity of functions and protein scaffolds that were hidden within those venoms; they unveiling a huge amount of intraspecific variation with ontogenetic and geographical bases
[1]. Nevertheless, most of snake diversity lies within a group commonly known as colubrids (Superfamily Colubroidea sensu Zaher et al.
[9]), that have been out of the spotlight of toxinological studies for most of history
[8–11].
In Colombia, colubrids account for ⁓70% of the ⁓340 currently recognized snake species; and they are distributed in 60 genera
[8]. These snakes possess a wide spectrum of morphological and physiological traits that separates them from front-fanged snakes (i.e., vipers and coralsnakes)
[12–14]. Colubrids are endowed with a secretory maxillary gland, called Duvernoy’s gland, that is homologous (similar in position, structure, and evolutionary origin) to the venom gland of vipers and elapids
[13–15]. This gland produces toxins, and it is connected to one of the two different types of maxillary dentition present in colubrids. The first type of dentition consists of rows of undifferentiated teeth with an absence of grooved fangs, known as aglyphous. The second is characterized by the presence of posteriorly enlarged grooved fangs, known as opisthoglyphous
[13–17].
These characteristics granted them different mechanisms of subduing prey including constriction, venom inoculation, or a mixture of both methods. In addition, colubrids exhibit a wide spectrum of ecological specializations that allow them to adapt to a wide variety of habitats, diets, and behaviors
[12,18–24]. While most colubrids are terrestrial or arboreal, feeding mostly on birds, mammals, amphibians, or other reptiles, some species are specialized to thrive in aquatic or semifossorial environments, changing their preferred prey to fish and invertebrate animals
[16,19,22,24–27].
Colubrids exhibit a myriad of ecological specializations that are accompanied by similarly diverse changes in the function of their venoms. Some species possess venoms dominated by proteolytic enzymes (enzymes that break down proteins) similar to those found in vipers; others emulate the neurotoxic profile common in elapids; and some groups even have lost their Duvernoy’s gland altogether
[1,11,15,16,28–33].
Notably, the venom inoculation system of colubrids tends to be less efficient than that of viperids and elapids, mainly because their Duvernoy’s gland does not possess the insertion of a compressor muscle that can generate the pressure needed to efficiently penetrate the tissue of their prey
[1,15,16]. The lack of an efficient venom delivery system is one of the main reasons that cause them to be considered less medically relevant. Thus, their venoms have been poorly studied in comparison to their viperid and elapid counterparts
[10,11,34]. This medical bias transformed these species into a neglected group within toxinological sciences worldwide, and Colombian species are no exception.
Although several colubrids possess venom, only a small number of genera distributed in Colombia have been reported to cause severe or mild envenomation in humans
[35]. However, accidents with some species might be far from harmless and may be underestimated in epidemiological studies done in the country
[17,36] (see Chapter 9). Moreover, the dearth of information about the real number of yearly accidents involving colubrids increases the already existing bias against understanding the biological activities and composition of their venoms
[10].
This bias is not only caused by the relatively low medical significance they have, but also by several factors that make the research of colubrid venoms an arduous task. For example, the difficulty of efficiently maintaining these snakes in captivity and extracting venom for research purposes, the low venom yields, the scarce chance of encountering venom-producing rear-fanged species, the low toxicity for humans and in murine models, the high prey specificity of their toxins, and the slow spread of their venom when injected into tissues are some of the main reasons that slow venom studies of colubrid species
[37–41].
Most Colombian colubrids lack data about the composition, pharmacological effects, or biological function of their venom or oral secretions. Fortunately, recent studies performed with Colombian and non-Colombian species have partially unveiled the enormous diversity of toxins that are present in the venoms of this group of snakes
[11,32–34,42]. Complex mixtures of proteins, like those found in vipers and elapids with myriad enzymatic and non-enzymatic activities have been reported in various venoms of colubrid species, turning them into an interesting prospect for searching for new, previously undescribed toxins.
Functional studies have revealed that some venoms from colubrids show similar potencies to those from vipers. Bites from these colubrid species can even induce serious clinical manifestations that might incapacitate a patient
[17,36,43–51]. In this chapter, we will review the available information about the composition, function, and clinical manifestations of the venom of colubrids distributed in Colombia, focusing on the untapped potential that lies in this group and the future of toxinological studies in the Colombian panorama.
2. The venom of “colubrid” snakes
Most snake diversity in Colombia is contained within a single group of opisthoglyphous and aglyphous species (see Chapter 1) known as the superfamily Colubroidea
[8]. Currently, there are three recognized subfamilies inhabiting Colombia: Colubrinae (~50 species in ~15 genera), Dipsadinae (~187 species in ~44 genera) and Sybinophiinae (one species in one genus). Venom-related information is available only for some genera of Colubrinae and Dipsadinae, focusing on compositional and functional data
[11,28,29,32–34,42,52–57]. In this section we shall discuss the trends observed in the venoms of both subfamilies of Colombian colubrids.
2.1. Subfamily Colubrinae
Historically, most genera and species from this subfamily lack any type of venom-related information
[10,11]. Recently there has been a continuous increase in the amount of published data tackling colubrine venoms. Several works including Asian, African and American species have unveiled a large repertoire of functionally diverse toxins
[11,28,29,52,55,58–61].
Some species from this group resembling viper venoms (see Chapter 3) possess a venom profile dominated by snake venom metalloproteinases (SVMPs), a group of enzymes that break down tissue and cause hemorrhage and other local manifestations
[58,61]. However, most of the toxin diversity found in this subfamily is composed of non-enzymatic post-synaptic neurotoxins known as three-finger toxins (3FTxs)
[28,52,55,59,60]. These proteins are common components of elapid venoms (see Chapter 5), causing a potent neuromuscular blockade that leads to asphyxiation by respiratory arrest and a rapid death
[3,62]. Interestingly, 3FTxs of Colubrinae appear to have evolved to become prey-specific, being more toxic towards certain types of prey
[63]. Due to the broad spectrum of prey items that Colubrinae species with 3FTxs feed upon, we can only hypothesize about the different amount of specific functional motifs that are hidden in their neurotoxins.
In Colombia, all species with venom-related data have a noticeable neurotoxic profile with a rather simple compositional pattern in terms of the number of toxin classes. Genera
Spilotes,
Leptophis,
Tantilla,
Rhinobothryum and
Oxybelis contain the species with available compositional information
[29,30,52,55,60]. Their venoms are dominated by neurotoxic 3FTxs and cysteine-rich secretory proteins (CRISPs,
see Chapter 5). The latter are a widespread class of toxins commonly seen in most member of the Colubridae that are known to interfere with smooth muscle contraction by blocking ion channels
[1,64,65].
In addition, other minor components reported in most studied genera are SVMPs and C-type lectins (CTLs) that are associated with local effects such as hemorrhage, edema, necrosis, and platelet-related hemostatic disorders
[66]. This compositional information corresponds with the low hemorrhagic and myonecrotic activities observed in most Colombian Colubrinae
[30], although those studies were carried out with venom samples from foreign populations. Intraspecific geographic variation of colubrid venoms is still poorly explored and an unresolved research area for Colombian populations (see Chapter 5).
Interestingly, the Duvernoy’s gland is not the only toxin-producing cephalic gland found in Colubrinae species. A recent study with a species of
Tretanorhinus, revealed that the Harderian gland is also capable of producing highly expressed venom-like transcripts
[57]. This gland is located behind the eye of the snake and is proximal to the Duvernoy’s gland. It is thought to be closely related to the lacrimal gland aiding in the lubrication of the eye, but it has been proposed to fulfill several other roles such as acting as an accessory salivary gland that may secrete digestive enzymes or provide a source of lubrication
[67,68].
The expression profile of the Harderian gland of
Tretanorhinus variabilis revealed a series of putative toxins including P-III SVMPs, CTLs, and CRiSPs that are three of the main toxins reported in other Colubrinae
[57]. These findings indicate that the production of toxin-like proteins is not limited to the Duvernoy’s gland in colubrids, but it is not clear if these other oral glands that are producing putative toxins represent a trophic advantage (ecological function).
Overall, this subfamily contains species with prey-specific highly neurotoxic venoms dominated by 3FTxs and CRISPs. The molecular targets, biochemical characterization, and function of these colubrid toxins remain poorly studied. Therefore, they represent an interesting model to explore novel molecular mechanisms that might be pharmacologically exploitable since they interact with several ion channels that modulate nerve responses
[62,65]. Nevertheless, most species and genera distributed within Colombian territory lack compositional data that is needed to gain a better understanding of the evolutionary processes that might be shaping venom composition and function.
2.2. Subfamily Dipsadinae
Contrasting with the neurotoxin dominated venoms that are widespread across the Colubrinae, this subfamily has a more proteolytic venom profile with some exceptions (e.g. genus
Helicops)
[11,42]. The subfamily Dipsadinae contains some genera that are known to be medically important and cause serious cases of envenomation with a predominance of local symptoms (i.e., hemorrhage, edema, necrosis)
[31–33,56,69–71]. Compositional studies performed on Colombian genera revealed that Dipsadinae venoms are extensively dominated by proteolytic P-III SVMPs and CTLs that directly correlate with most envenomation clinical symptoms informed in the case reports
[11,32–34,42].
However, a recent study found that three tribes of this subfamily (Tachymenini, Xenodontini and Conophiini) had a different type of metalloproteinase as the main enzymatic component of their venoms
[42,53]. These enzymes were identified as snake venom matrix-metalloproteinases (svMMPs) and represent three independent events of gene recruitment in the evolutionary history of this group of snakes (See section 4 on this chapter).
Moreover, the tribe Hydropsini possesses a compositional pattern that is neither SVMP nor svMMP dominated, agreeing with the neurotoxic venom profile reported in
Helicops angulatus and the CTL-dominated profiles observed in other species of the genus
[42,72,73]. So, we can separate three big compositionally different groups of Dipsadinae venoms:(1) genera with venoms dominated by P-III SVMPs (e.g.
Leptodeira,
Pseudoboa,
Philodryas and
Oxyrhopus)
[11,32,34,54]; (2) genera with venoms dominated by svMMPs (e.g.
Erythrolamprus,
Lygophis and
Thamnodynastes)
[11,42,53]; and (3) genera with a unique profile with and abundance of CRISPs and CTLs, similar to what is found in some Colubrinae species (e.g.
Helicops)
[42,72,73].
Functional analyses performed in species of
Philodryas,
Leptodeira,
Thamnodynastes,
Erythrolamprus,and
Pseudoboa verified the proteolytic effects predicted by the analyses of venom composition. Most species of dipsadids show strong proteolytic capabilities
in vitro, even comparable to those of some
Bothrops species
[32–34,42,56].
Interestingly, in some species of the Pseudoboini tribe (Genera
Clelia and
Boiruna), a new type of phospholipase A2 (PLA2) has been detected as a dominant toxin
[11,61,73]. These PLA2s are different from those found in vipers and elapids (see Chapter 5) as they belong to the IIE group, typically found in brain, heart and uterine tissue of mammals
[1,74]. Recently, a transcriptomic and proteomic analysis of the whole tribe revealed that these PLA2-IIE that dominate the venom of these genera possess a shorter C-terminal tail when compared to the endogenous protein that fulfills physiological functions in other snakes
[73]. This shortening could be a convergent process to what is observed in other enzymatic toxins that have evolved into more compact scaffolds from a larger endogenous gene
[73].
Although there is still little information available about these toxins, they are likely secretory, calcium-dependent enzymes that primarily target extracellular phospholipids from their primary and tertiary structures, that resemble the ones observed in PLA2s from vipers
[73]. Moreover, enzymatic assays with venoms of the Pseudoboini tribe confirmed that they are catalytically active, with
in vitro activity levels similar to those observed in viper venoms rich in PLA2s
[1,73,74]. This PLA2 dominance is unique amongst colubrids so far, and represents the fifth recruitment of the PLA2 scaffold to snake venoms, showing that venom tends to converge towards the same optimal toxin scaffolds across different snake families
[73].
Additionally, an acidic lipase was reported as a new putative toxin in the venom of
Phalotris mertensi (False coral) that might represent the recruitment of a new toxin scaffold to the venom repertoire of colubrids
[74]. Overall, studies of colubrid venoms have resulted in the discovery of several novel scaffolds of toxins that in some cases lead to phenotypical convergence with front-fanged snakes and in others they result in new, unique venom components
[11,42,61,75].
2.3. Compositional trends of colubrid venoms
Comparing the general trends observed in the venom compositions of colubrids of both transcriptome and proteome, some interesting patterns appear (Figures 1 and 2). The overall dominance of proteolytic venoms in the Dipsadinae subfamily is clear where two distinct types of zinc-dependent metalloproteinases appear to have been recruited into the venom arsenal of this subfamily (Figure 1)
[42]. Snake venom metalloproteinases are present across several groups of advanced snakes (see Chapters
3 and
5) and are likely to have been ancestrally recruited before the separation of vipers and all other venomous snakes
[11,76–79].
On the other hand, some tribes (Conophiini, Tachymenini, and Xenodontini) of the Dipsadinae subfamily possess another type of proteolytic enzyme (svMMPs) that dominates their venoms. These proteins are likely to have been recently recruited into the venom glands of species of Dipsadinae and are the dominant toxin class in some groups
[42].
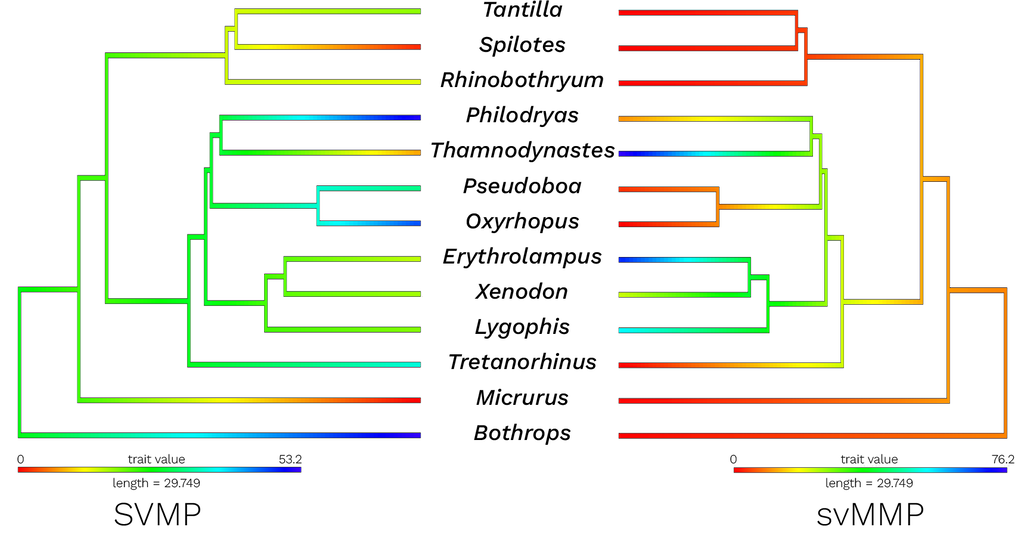
Figure 1. Ancestral state reconstruction of compositional data for SVMPs (left) and svMMPs (right). Red color indicates lower proportions of that toxin while blue colors indicate higher proportions of that toxin; yellow to green color scale depicts moderate proportions. Data for
Rhinobothryum were obtained from proteomic analyses
[55] and data for
Tretanorhinus belong to a study that analyzed the Harderian gland
[57].
Micrurus and
Bothrops were used as outgroups
[80,81]. Branch lengths were obtained from Zaher et al.
[9].
Venoms of Colubrinae species show highly neurotoxic venoms dominated by 3FTxs that are post-synaptic toxins that cause neuromuscular blockades by acting at the motor endplate (Figure 2)
[62]. This compositional pattern resembles the one observed in some coralsnakes and marks a conspicuous difference between Colubrinae and Dipsadinae venoms. Nevertheless, some species of Dipsadinae possess neurotoxic effects, but these are thought to be mediated by CRISPs or to be secondary effects of the proteolytic action of their toxins
[56,72].
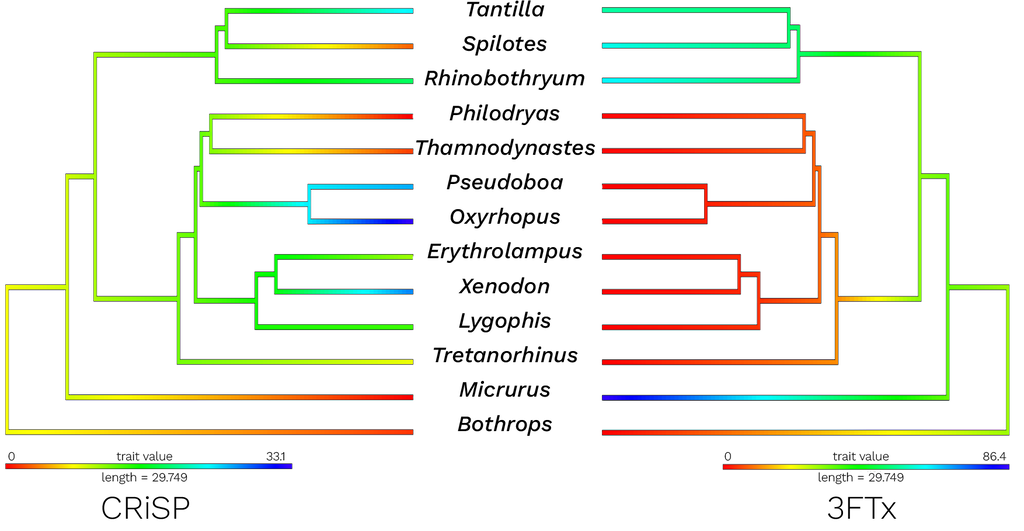
Figure 2. Ancestral state reconstruction of compositional data for CRISPs (left) and 3FTxs (right). The red color indicates lower proportions of that toxin while blue colors indicate higher proportions of that toxin. Yellow to green color scale depicts moderate proportions. Data for
Rhinobothryum were obtained from proteomic analyses
[55] and data for
Tretanorhinus belong to a study that analyzed the Harderian gland
[57].
Micrurus and
Bothrops were used as outgroups
[80,81]. Branch lengths were obtained from Zaher et al
[9].
Overall, venoms from colubrid species possess a diversity of toxins similar to that of vipers and elapids (see Chapters
3 and
5) with some groups favoring 3FTx-dominated highly neurotoxic venoms and others SVMP/svMMP-dominated highly proteolytic venoms (Figure 3). Moreover, even though the vast majority of colubrid species lack any kind of venom-related information, an enormous diversity of toxin scaffolds and functions unique to this group of snakes has already been unveiled
[11,37,42]. These convergent and divergent patterns indicate that changes in venom composition could be linked to both phylogenetic inertia and ecological factors leading to convergence
[42,82,83].
This duality has already been noted in the past for the variation of the activity of two enzymatic toxins from Australian elapid species. Although enzymatic activity showed a stronger correlation to phylogeny in most genera, divergent activity levels were reported for two closely related species of
Denisonia that are frog specialists
[82]. Therefore, there might be unknown underlying factors shaping venom composition and function in front-fanged, rear-fanged, and even aglyphous snake species. Studies that link ecological variables to compositional and functional venom data, along with phylogenomic information, are needed in order to gain a better understanding of the evolutionary processes that mold the venom phenotype of extant snakes, especially in colubrid species.
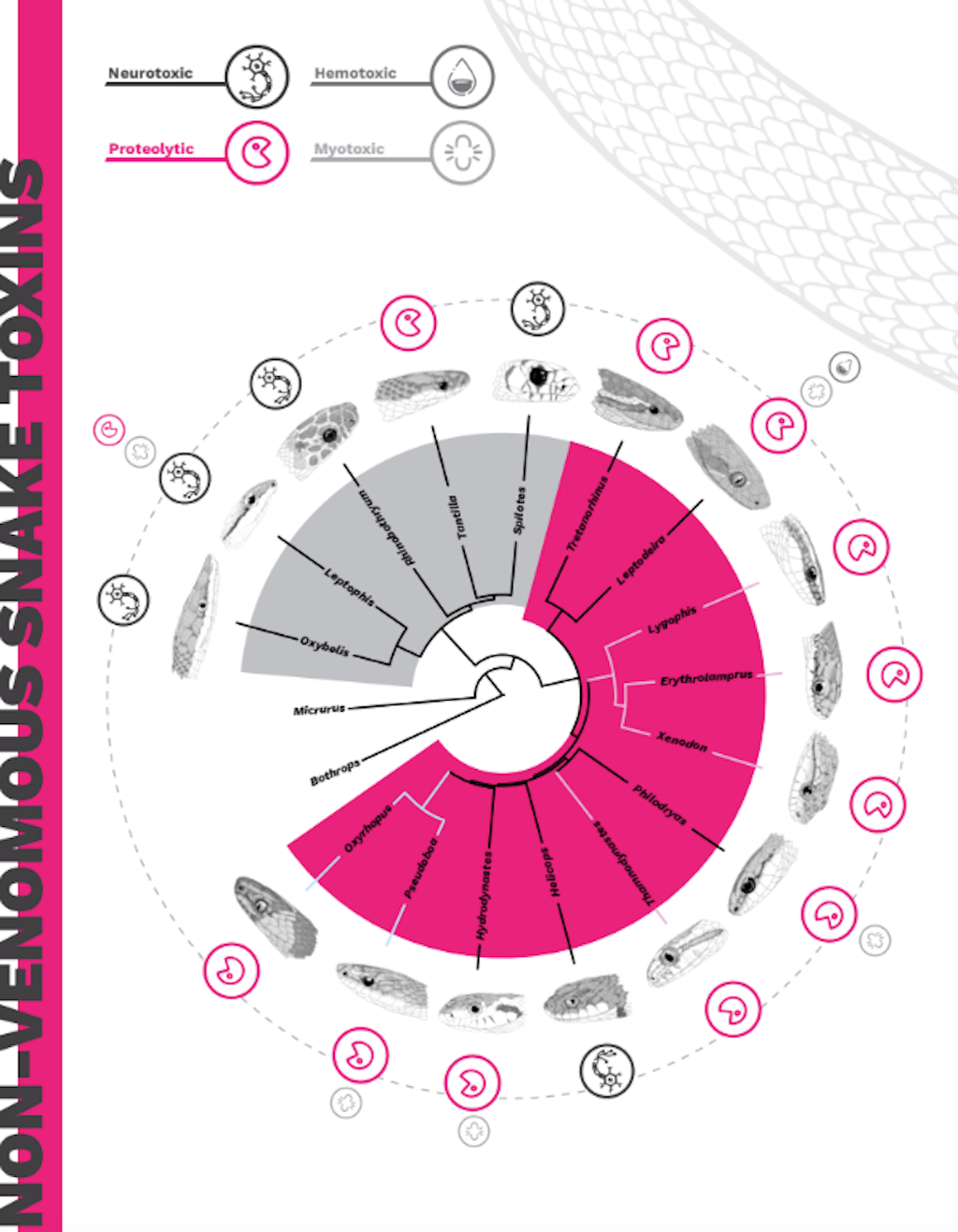
Figure 3. General overview of the known venom profiles of colubrids.
2.4 Venomic notes of Colombian colubrids
Envenomation by colubrid snakes in Colombia is a poorly known phenomenon, consequently, the clinical relevance of these species in snakebite accidents has been historically underestimated
[106]. This does not mean that envenomation by colubrid snakes do not occur. In fact, during the last decade a total 347 cases were notified in Colombia (see Chapter 9), and those caused by
Leptodeira annulata and
Erythrolamprus bizona were documented with detail
[106,107]. However, the true snakebite incidence caused by colubrid snakes is masked. Following, we present the available venomic information of the colubrid snakes (aglyphous and opisthoglyphous) with potential medical importance in Colombia.
Genus Erythrolampus.— Recently, it was found that venoms from this genus are mainly dominated by a novel proteolytic scaffold known as snake venom matrix metalloproteinases (svMMPs) and a few other minor toxins such as SVMPs, CTLs and CRISPs
[42]. Functionally, the venom of Colombian populations of
Erythrolampus bizona produces partial neuromuscular blockade causing notable morphological alterations with higher doses involving disorganization of muscle fascicles, interstitial edema, and expressive muscle fibers necrosis
[56]. The neuromuscular effects observed seem to be the result of the high proteolytic activity rather than a neurotoxic mechanism which fits perfectly with the mainly proteolytic profile of the reported venom proteins
[56].
Leptodeira annulata species complex.— This species is known for its harmless and inoffensive behavior
[108] (Figure 4). Nevertheless, there are reported cases of moderate local envenoming when its bite is provoked due to improper handling
[106,109]. Venom of Colombian populations from the Magdalena Valley has been explored
[33,34], finding protein bands with molecular masses from ~15 kDa to ~140 kDa. The most conspicuous bands were seen at ~35 kDa, ~40 kDa and ~50 kDa, as well as a prominent band at the migratory front (~15 kDa). In addition, chromatographic analysis (RP-HPLC) showed a total of 38 separate peaks, indicating a highly complex venom composition, which can be equal or superior that observed the venom of some
Bothrops species
[34].
Recently, a complete toxinological characterization of venom from
L. annulata was reported. The venom had no esterase, L-amino acid oxidase or thrombin-like activity, but was proteolytic active towards several substrates including fibrin, fibrinogen, gelatin and casein. The proteolytic activity is probably mediated by metalloproteinases as it was only inhibited by metal ion chelators (EDTA and phenanthroline)
[34]. Besides, the venom did not aggregate rat platelets or inhibit collagen-induced aggregation, but partially inhibited thrombin-induced aggregation. The venom was also hemorrhagic (inhibited by EDTA) and increased the vascular permeability in rat dorsal skin. Additionally, the venom produced myonecrosis and increased serum creatine kinase concentrations in the gastrocnemius muscle
[34].
Leptodeira annulata venom preparations applied to nerve-muscle, produce a moderate neuromuscular blockade without causing a complete blockade. Moreover, it significantly inhibits the contracture response to exogenous potassium chloride. In contrast, in mice nerve-muscle preparations, the venom of
L. annulata does not cause significant neuromuscular blockades. However, it induced minor muscle membrane depolarization without affecting the postsynaptic receptors
[33]. These neuromuscular effects observed are the result of an unspecific proteolytic activity of the venom rather than a neurotoxic mechanism. On the other hand,
L. annulata venom causes clear histological alterations in both chick and mice models, including swollen fibers and necrosis. These effects seem to be related to the high proteolytic and mild PLA2 activity exerted by the venom
[33].
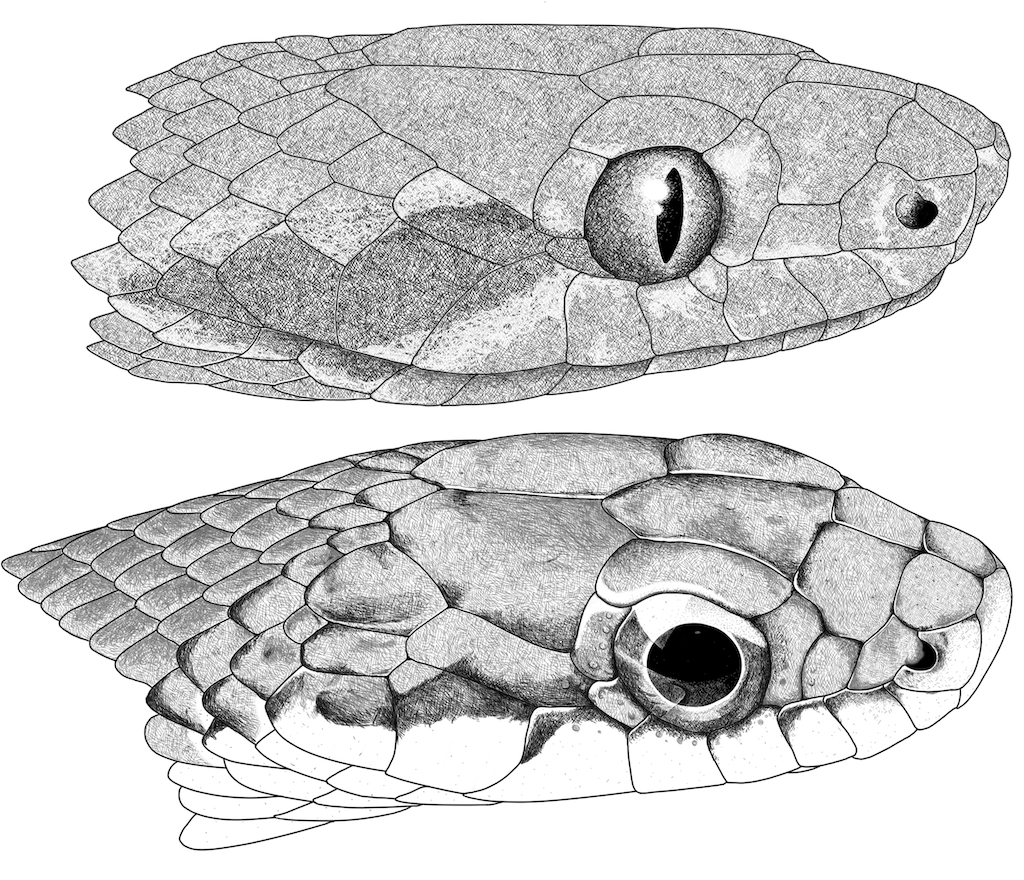
Figure 4. Head illustrations of
Leptodeira annulata and
Leptophis ahaetulla species complex. (Top) head in lateral view of
Leptodeira annulata from Pacurita, Quibdó, Chocó, Colombia (JDL 31754). (Bottom) Head in lateral view of
Leptophis ahaetulla from Caño Cristales, Riosucio, Chocó, Colombia (IAvH-R1458). Illustrations by Oscar A. Ramírez Ruiz.
Genus
Leptophis.— Among the
Leptophis species inhabiting Colombia, only
Leptophis ahaetulla has venom studies. Although it is a commonly found genus across its distribution range, there are few studies regarding venom composition or function with Colombian populations (Figure 4). Rodríguez-González and Portillo-Portillo
[110] made the first work testing the toxic capacity of
L. ahaetulla venom. They tested the neurotoxic effect of this species venom in the sciatic nerve of the frog
Rhinella horribilis. However, the venom did not disturb the action potential of the nerve, and the author hypothesized that it had a minor biological function.
Recently, Sanchez et al.
[30] performed a functional and compositional characterization of venom from an Argentinian population of
L. ahaetulla revealing a compositional profile dominated by small proteins (<10.0-30.0 kDa) identified as CTLs, CRISPs and 3FTxs. Consequently, proteolytic and myotoxic activities of the venom from
L. ahaetulla were practically null, as no enzymatic protein was identified as an important component of the venom. Moreover, lethality tests in mice showed no signs of lethal effects in doses as high as 20 µg/g, although this might be explained by prey-specific toxicity (frogs and lizards) as has been observed in other colubrid with neurotoxic venoms
[60,63,111]. Nevertheless, studies about functional and compositional characterization of
L. ahaetulla venom from Colombian populations have not been performed yet.
Genus
Lygophis.— There is not much information regarding the venom composition and function of
Lygophis snakes. A recent work analyzing the venom gland transcriptome of
L. flavifrenatus from Brazil found that its venom shows a highly proteolytic profile following the trend observed in other closely related genera (e.g.,
Erythrolamprus), with svMMPs as the main type of proteolytic enzymes among the other minor venom components
[42].
Genus
Oxybelis.— A few human accidents involving
Oxybelis species have been reported across its known distribution
[112–114]. Generally,
Oxybelis envenomation consists of local symptoms like pain, erythema, and hemorrhage, rarely accompanied by tachycardia and dizziness (Figure 5).
Regarding its venom, in the last decades some interesting research has been published. The venom from the green vine snake (O. fulgidus) seems to be compositionally simple with venom proteins within the 6-9 kDa, 25 kDa and 50-66 kDa size ranges, highlighting the presence of 3FTx
[111]. Moreover, a neurotoxin from this venom, which is the most abundant component, was purified and characterized. Fulgimotoxin is a 3FTx with very high lethal toxicity towards native lizards (Anolis species were more sensitive than
Hemidactylus species) and little to no effects on mammals (mice)
[111].
Recently, the venom proteome from two species was described. The venom proteomes were of low compositional complexity with four detected protein families (LAAO, P-III SVMPs, CRISP and 3FTx)
[60]. In general terms, both venoms are highly neurotoxic with lethal activity in lizards, low proteolytic activity, and lacking fibrinogenolytic activity
[60].
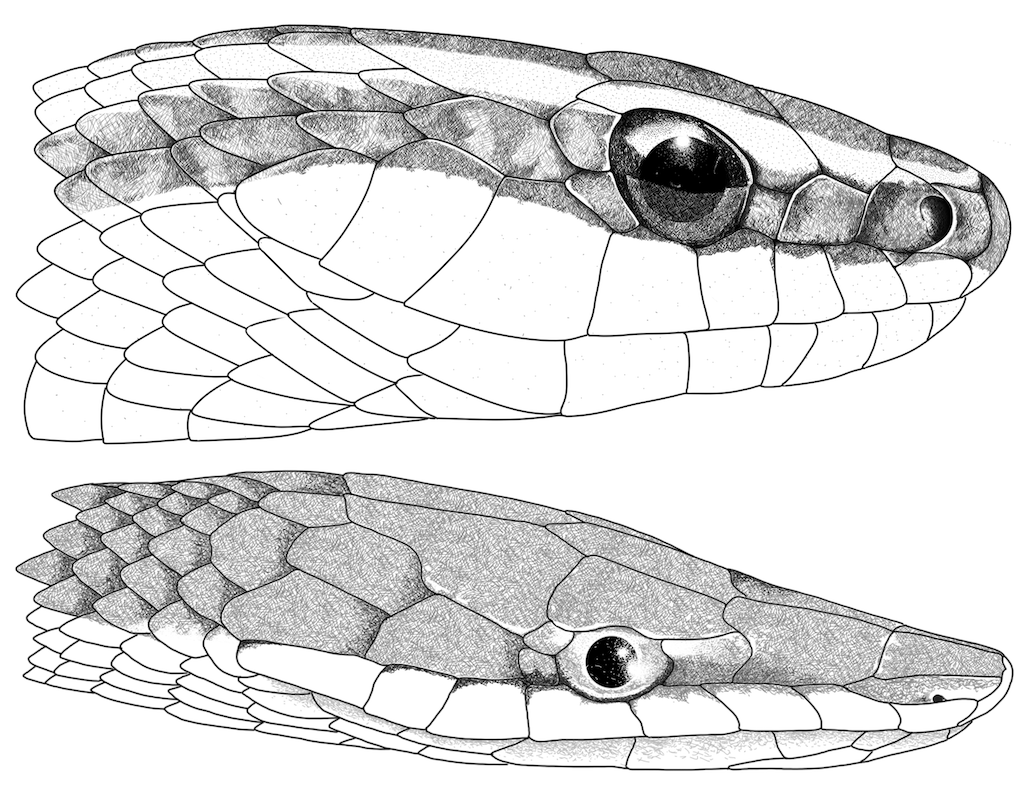
Figure 5. Head illustrations of
Lygophis
lineatus and
Oxybelis aeneus. (Top) head in lateral view of
Lygophis
lineatus from Universidad Nacional de Colombia sede de La Paz, La Paz, Cesar, Colombia (TAS 2576). (Bottom) Head in lateral view of
Oxybelis aeneus from Riosucio, Chocó, Colombia (IAvH-R1458). Illustrations by Oscar A. Ramírez Ruiz.
Genus
Oxyrhopus.— Unfortunately, there is not much information available about the venom composition or function of this group of snakes. However, the little data that exists suggests that venoms of
Oxyrhopus species are dominated by SVMPs, CRISPs and CTLs, following the same trend observed in most of Dipsadinae
[11]. Nevertheless, other venom components include PLA2s-IIE which are of special interest because it represents a class of PLA2 different than that found in vipers and elapids, perhaps being a new event of toxin recruitment into the venom gland arsenal of this group of snakes as was recently observed in other species of the Peudoboini tribe
[11,73]
Genus
Philodryas.— Among non-front fanged colubrid envenoming in South America (Figure 6), Philodryas species cause the most cases reported
[17,84,115–118]. Thus, they are the colubrid snakes with the most comprehensive venom-related data compared to other colubrid genera
[54,119,120]. Particularly, due to the wide distribution of
P. olfersii, its venom has been widely studied. This venom is highly proteolytic, produces edema, hemorrhage and myonecrosis in animal models
[121–124]. Some components of the venom have been isolated: a myotoxin
[125], four metalloproteinases and one serine protease
[126]. The characterization and identification of these venom components agree with the proteomic and transcriptomic analyses performed on this species venom
[54,127]. Functionally, the venom of
P. olfersii degrades fibrinogen and fibrin, and it is devoid of thrombin-like activity, platelet-aggregating inhibitory effects
[123]. The venom also does not produce hematological alterations having limited effects on circulating cytokine concentrations
[128]. Although different effects are observed
in vivo using murine models, primary manifestations in human envenoming are local effects such as pain, edema, erythema and ecchymosis
[85]. Currently, no cases of envenoming by species of the
Philodryas genus have been reported in Colombia.
Genus
Rhinobothryum.— Field encounters with these species are usually scarce, thus,
Rhinobothryum venoms are poorly known (Figure 6). Recently, a study with
R. bovallii revealed that it possesses a neurotoxic venom as observed in other genera of the colubrinae subfamily (e.g.,
Spilotes,
Leptophis and
Oxybelis)
[55]. Proteomic analyses revealed a rather simple compositional profile dominated by neurotoxic CRISPs (8.2%) and 3FTxs (86.5%) with a small proportion of proteolytic SVMPs (2.4%). Taxon-specific neurotoxins have been repeatedly reported in venoms of colubrids with specialized diets mainly focusing on birds and/or lizards. However,
Rhinobothryum species have a very diverse diet, so functional studies aiming to uncover the specificity of their neurotoxins are needed to gain a better understanding on toxin evolution and venom resistance in those prey models. Currently, cases of envenomation by
Rhinobothryum species have not been reported in Colombia.
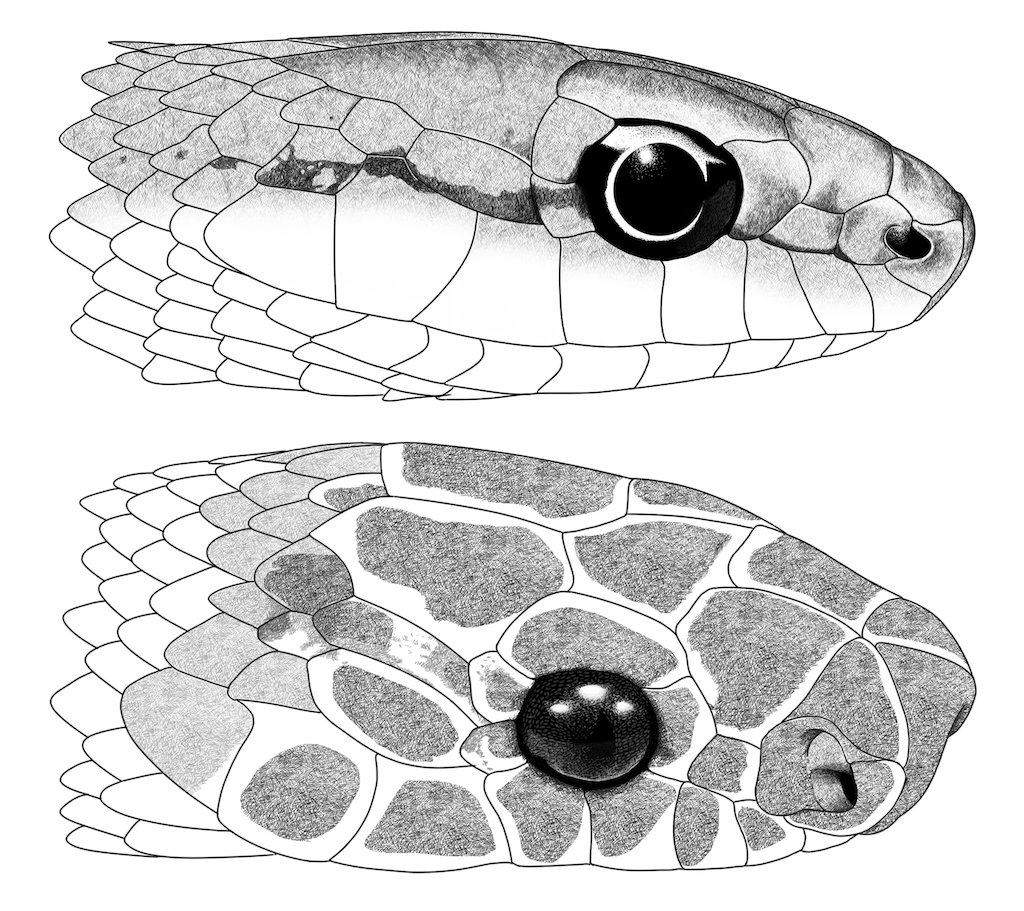
Figure 6. Head illustrations of
Philodryas
olfersii and
Rhinobothryum
bovallii. (Top) head in lateral view of
Philodryas
olfersii from Puerto Rondón, Arauca, Colombia (IAvH-R2878). (Bottom) Head in lateral view of
Rhinobothryum
bovallii from Pedernales, Manabí, Ecuador (QCAZR -R5757). Illustrations by Oscar A. Ramírez Ruiz.
Pseudoboa neuwiedii.— Venom of
P. neuwiedii from Tolima was biochemically characterized by Torres-Bonilla et al
[129]. In their analyses they report its SDS-PAGE profile and the identification of its components by LC-MS/MS. Venom from this species shows protein bands ranging from ~15 kDa to ~200 kDa, with the most conspicuous bands between ~50-75 kDa and ~20-25 kDa. These bands were identified as SVMPs and CRISPs, resembling the reported compositional pattern observed in other closely related species at the transcriptomic level
[11]. Among components identified, less intense bands were CTLs, PLA2s and more notably, svMMPs, a scarcely studied family of proteinases recently reported as a major component of the venoms of some dipsadids
[129].
Moreover, venom from
P. neuwiedii produces moderate neuromuscular blockade (50% blockade after ⁓80 min) with significant reduction of the contracture responses to exogenous potassium chloride and acetylcholine after 120 min of incubation. The venom caused extensive histopathological alterations in muscle preparations. The muscle fibers lost their normal and typical polygonal profile with swollen or vacuolated fibers leading to failure of permeability regulation. The neuromuscular effects observed are the result of a general proteolytic activity rather than a neurotoxic mechanism
[56].
Pseudoboa neuwiedii venom is highly proteolytic towards azocasein with optimal temperature of 37 °C, pH 8.0, extremely active in gelatin and displays moderate PLA2 activity. The venom also degrades the Aα chain of fibrinogen in vitro, lacks coagulant activity and delays the coagulation time during plasma recalcification
[129].
Genus
Spilotes.— Currently, there are no reports of envenomation cases or characterized venom properties from this species. However, there is recent research addressing the venom evolution of another species of the genus (S. sulphureus). The venom from
S. sulphureus was analyzed combining specific prey assays with venom gene expression and venom phenotype
[63]. Two novel neurotoxins (3FTx) were identified: sulditoxin and sulmotoxin 1, which are the most abundant venom proteins with opposite prey-specific toxicities. Sulditoxin is highly toxic to lizards and non-toxic to mammals, whereas sulmotoxin 1 exhibits the reverse trend
[63]. This is also the first case in which a bimodal and opposite pattern of toxicity for toxins is shown for a venom in relation to diet
[63].
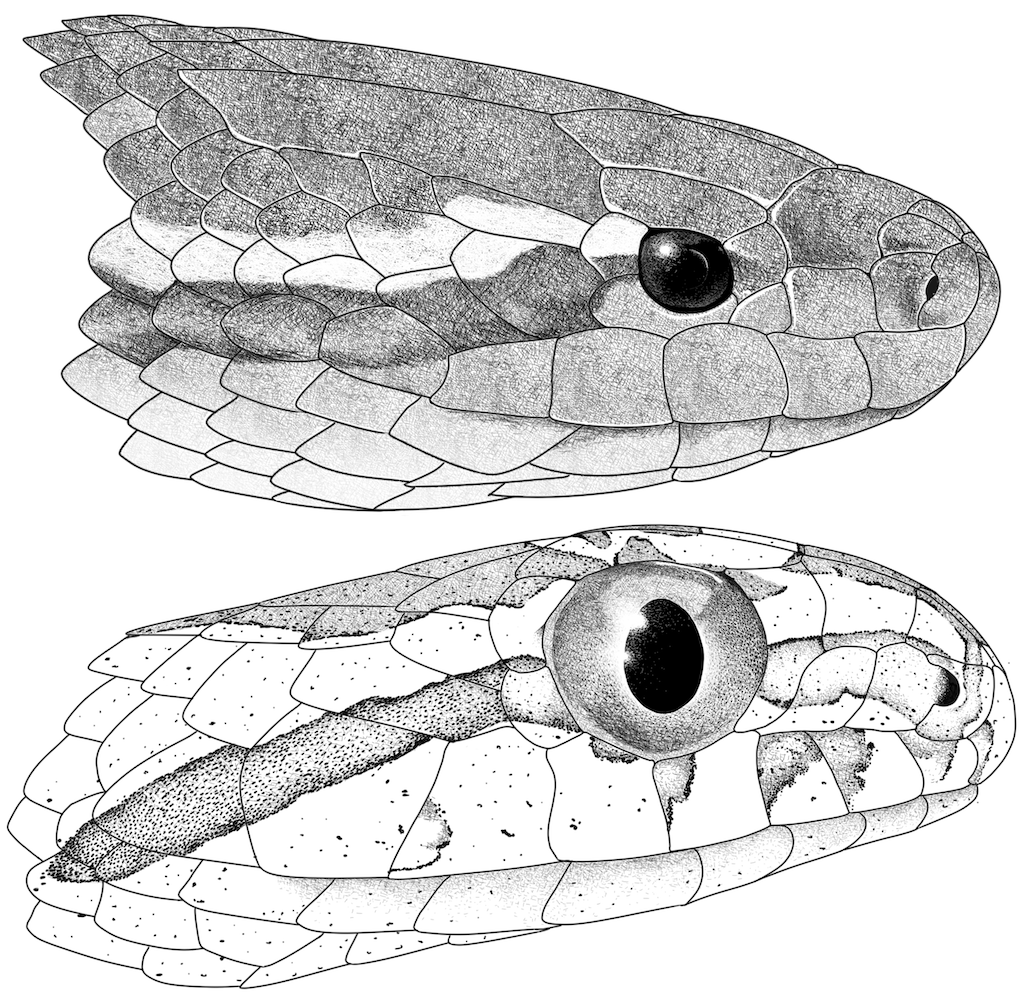
Figure 7. Head illustrations of
Tretanorhinus nigroluteus and
Thamnodynastes dixoni. (Top) Head in lateral view of
Tretanorhinus nigroluteus from vereda Cacarica, Riosucio, Chocó, Colombia (IAvH-R 2002). (Bottom) Head in lateral view of
Thamnodynastes dixoni from Puerto Carreño, Vichada, Colombia (JDL 31383). Illustrations by Oscar A. Ramírez Ruiz.
Genus
Tantilla.— Despite the enormous species richness within the
Tantilla genus, only
T. nigriceps, a species distributed in southern North America, has venom studies. These studies highlight that this genus possesses a venom highly neurotoxic. However, their small size and very low venom yields make them harmless to humans
[37,38]. The venom gland transcriptome of
T. nigriceps was dominated by neurotoxic 3FTxs (54%) and CRiSPs (24%), with smaller proportions of SVMPs (18%) and CTLs (3%)
[52]. The highly specialized diet of
Tantilla species would make their neurotoxins an amazing model to study toxin specificity and evolution.
Genus
Thamnodynastes.— Snake species of the genus
Thamnodynastes are notorious for their reported envenoming cases which generally include symptoms like edema, radiating pain, ecchymosis and hemorrhage
[46,130,131] (Figure 7). Nevertheless, no venom related information was available until 2012 when Ching et al
[132] published the proteomic and transcriptomic characterization of the venom of
T. strigatus. Their findings revealed that a new type of metalloproteinases, called svMMPs, were the dominant toxin class in both the venom proteome and the venom gland transcriptome. svMMPs accounted for 46% of the whole transcriptome and 76% of all the annotated toxins which included SVMPs, CTLs, 3FTxs, CRISPs and other minor toxins. Recently, a transcriptomic screening including more species confirmed the dominance of svMMPs in the venom of this genus and showed that they maintain a highly proteolytic profile
[42].
Genus
Tretanorhinus.— Due to the scarce sampling and information readily available for this genus, there has not been a single study regarding venom composition for either of the two Colombian species (Figure 7). Nevertheless, the transcriptome of the harderian gland from a Cuban swamp snake
T. variabilis has been analyzed. This gland is located behind the eye of the snake and is proximal to the duvernoy’s gland. It is thought to be closely related to the lacrimal gland aiding in the lubrication of the eye, but it has been proposed to fulfill several other roles
[67,68,133]. These hypothetical functions include acting as an accessory salivary gland which may secrete digestive enzymes or provide a source of lubrication. In
T. variabilis the expression profile of the harderian gland revealed a series of putative toxins including P-III SVMPs, CTLs, and CRiSPs, which are three of the main toxins reported in other colubrids
[57]. These findings indicate that the production of toxin-like proteins is not limited to the duvernoy’s gland of colubrids, but it is not clear if these other oral glands that are producing putative toxins represent a trophic advantage.
3. Epidemiology and clinical manifestations
Envenomation by colubrid species has been greatly overlooked by epidemiological studies worldwide
[10,11,35,47,84]. This might be related to the fewer number of bites involving colubrids or underreporting, as well as the relatively minor clinical manifestations caused by their venoms
[35,43,45,47,49,85,86]. However, these mild effects could result in partial or full immobilization of affected limbs, causing a potential loss of productive work capacity for the patient (commonly a farmer or agricultural worker)
[17,36,44,48,51,85].
In Colombia the incidence and epidemiological records of snakebite as a public health metric is provided by the Surveillance System of the National Health Institute (Sistema Nacional de Vigilancia en Salud Pública: SIVIGILA, Spanish acronym) (see Chapter 9). The SIVIGILA publishes annual reports with the total number of snakebite cases including numerous amounts of data including information about the identification of the snake involved in the accident. However, the process of data collection and analysis involves the arduous revision of handwritten forms filled by medical practitioners from all around the country that had varying degrees of available taxonomical information. This complication is generally bypassed for commonly identified groups (e.g. genera
Bothrops,
Crotalus and
Micrurus), but it still is a difficult bias to overcome for most colubrid species that typically have various common names that change between different regions, and encompasses most of the snake diversity of the country
[8,10].
Therefore, the number of ambiguously identified or undetermined snakes that were reported is generally high with ~11,280 out of the ~49,143 cases between 2010-2020 (~20% of total cases). This is likely to increase the already existing identification bias that colubrid envenomation suffers, widening the gap of medical relevance between front-fanged and rear-fanged snakes
[10,11].
Fortunately, since 2012 the National Health Institute (Instituto Nacional de Salud: INS, Spanish acronym) has conducted several campaigns to educate about the prevention and handling of snakebite incidents, helping medical professionals to gain a better insight into snake identification. The positive impact of these initiatives becomes clear when analyzing the cumulative average of annual snakebites that were not identified in the country (Figure 8).
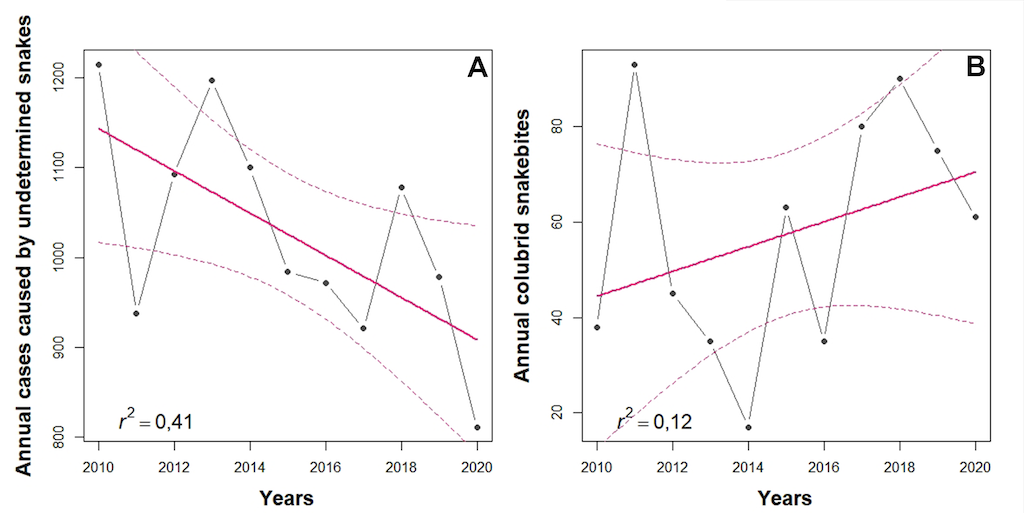
Figure 8. Linear regression model of the cumulative average of annual undetermined snakebites (A) and the annual colubrid snakebites (B) reported by the SIVIGILA (2010-2020). Grey lines and dots represent the yearly variation of snakebite cases. The solid pink line represents the best fit of the linear regression model, and the dashed pink lines shows the 95% confidence interval for the model. r2 values are shown on the bottom left of each graph.
The number of undetermined cases shows a diminishing trend in the last ten years (F= 6.15, p= 0.035). Therefore, we could expect an increase in the number of identified colubrid snakebites that were previously hindered as undetermined cases. When analyzing the annual occurrences of colubrid snakebites we observe an increasing trend. However, this tendency is not significant due to the heterogeneity of the data (F = 1.19, p= 0.30). Nevertheless, as the number of undetermined cases decreases, we expect to see an increase in the number of reported snakebites involving colubrids.
Currently, there are ~30 confirmed annual snakebite cases caused by recognizable snake species belonging to the Colubridae or the Boidae families in the SIVIGILA database (~541 and ~91 respectively from 2010-2020). These cases have a rather consistent profile of clinical manifestations (Figure 9). Pain, local symptoms, edema, erythema, systemic symptoms, paresthesia, and general sickness are among the major reported symptoms. Moreover, when comparing these occurrences with the clinical profile of undetermined cases we see a similar profile that might relate to the likely high proportion of colubrid-related cases that are labeled as undetermined.
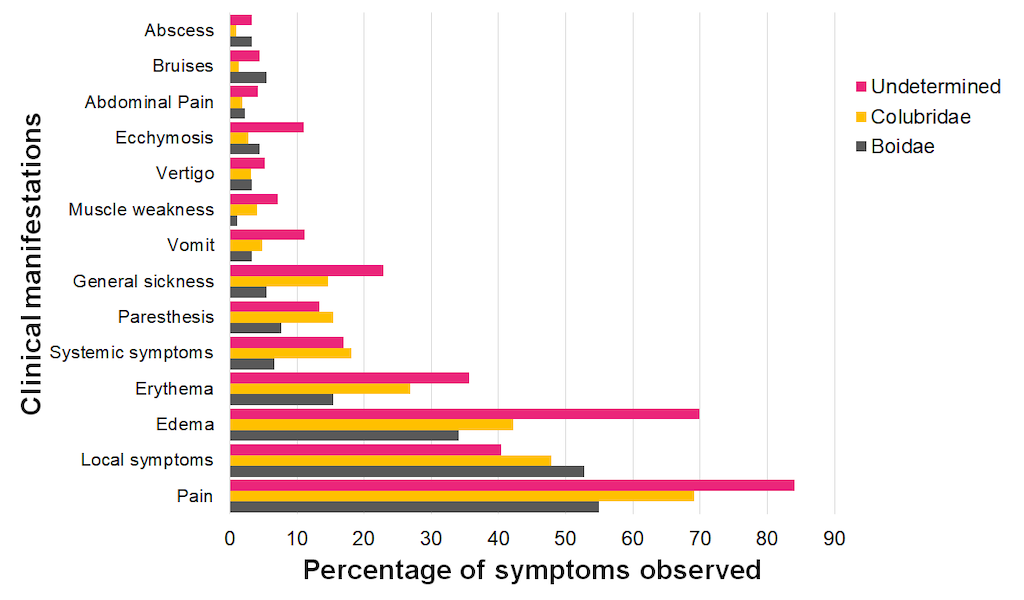
Interestingly, the SIVIGILA database reports the application of antivenom in some cases caused by either a Colubridae or a Boidae species (~159 and ~19 respectively). This is problematic as allergic reactions to polyvalent antivenom are known to cause medical complications
[87]. So, the correct and accurate identification of the species that caused the accident is a crucial stage to ensure the safety of the patient. Increasing the percentage of accurate identification of non-front-fanged snake species is key to obtain a complete panorama of the true epidemiological relevance of snakebites in the country.
Overall, the management of snakebite incidence in Colombia has improved in the last decade, although some challenges remain. There is an urgent need to increase the percentage of identified snakes involved in snakebite cases and to implement a more efficient system to compile and organize all the clinical information of each snakebite case. Moreover, colubrids and other non-front-fanged snake groups should be given a higher relevance in clinical studies and courses in order to decrease the number of undetermined snakebite incidents and ensure that each snakebite case is treated correctly.
4. The untapped biological potential of “colubrid” snakes
Historically, snake venoms have been a source of novel protein scaffolds with evolutionarily intriguing and pharmacologically applicable functions
[86–88] (see Chapter 10). However, as previously discussed, most research on venom diversity has focused on vipers and elapids, neglecting the majority of venom producing species
[11,39]. Fortunately, in recent years, there have been transcendent advances in our knowledge of colubrid venom composition that reveals a toxin diversity of similar magnitude to that of front-fanged snakes
[11,42,73]. In this section we will present the main direct and indirect applications of colubrid venoms and why they represent an interesting biological model for both evolutionary and pharmacological studies.
4.1 Colubrid venoms as a tool for understanding evolutionary processes
Snake venom is a trophic adaptation that is directly linked to the capacity for effective hunting and feeding, since it aids in paralyzing and killing prey
[63]. Therefore, venom composition and function are under heavy selective pressure and can only vary between a few optimal thresholds
[91]. Moreover, genes controlling toxins are generally recruited into the venom gland through a process known as gene duplication that allows for rapid diversification as new gene copies escape selective constraints, being free to mutate
[42,79,83,92].
Hence, snake venom toxins display a broad spectrum of diverse functions and molecular targets contained within few conserved structural scaffolds
[62,76,77,91,93]. These peculiarities have turned them into interesting models for studying phenotypical convergence, gene recruitment and co-option, predator-prey relationships, and several other evolutionary phenomena
[29,42,76,83,93].
Snake venoms are dominated by a relatively small number of protein families, with SVMPs, PLA2s, 3FTxs, and SVSPs as major components of a few optimal compositional profiles. However, most studies have few data from colubrid species; as they remain a sub-sampled group in venom-related research
[11,91]. Nevertheless, the few colubrid venoms that have been thoroughly studied have already revealed the occurrence of new toxin scaffolds that are likely to represent unique gene recruitment events
[42].
Recent studies of colubrid venoms have detected the occurrence of phenotypical convergence and recruitment of novel gene families in their venom repertoire. For example, snake venom matrix metalloproteinases (svMMPs) were recently found to be a major toxin class in three tribes of the dipsadinae subfamily, replacing SVMPs as the main proteolytic toxins in Xenodontini, Tachymenini and Conophiini
[42]. This toxin class was recruited independently in each of those tribes and represents a curious event of convergence among them. Interestingly, these newly reported proteolytic toxins evolved into simplified structures resembling the SVMPs of vipers, and they are thought to possess similar proteolytic capabilities
[42,53,76,77,94] (see Chapter 5). Examples of the maintenance of a phenotypic trait by recruiting new genes into a tissue are scarce
[42,83], but snake venoms, as a multi-factorial trait determined by countless proteins with overlapping and synergistic functions, compose a prime model for studying these intricate evolutionary events.
As a trophic adaptation, snake venom is a useful tool for understanding predator-prey relationships and species adaptations to changing environments. This is especially true for evolutionary hypotheses like “The red queen hypothesis” proposed by Leigh Van Valen, in which predator and prey are in a constant arms race to surpass one another
[95,96]. In this example, venomous snakes would evolve more specific and potent toxins towards their prey, while their prey would evolve resistance mechanisms or specific behaviors to avoid being preyed upon.
Several examples of toxin resistance/specificity have been reported for front-fanged snake venoms, involving both proteolytic and neurotoxic toxins. Venom resistance mechanisms are generally mediated by circulating serum factors or by adaptive changes in venom-targeted molecules
[97–99]. A classic example of the former are
α1B-glycoprotein homologs found in mongooses and opossums that inhibit either SVMPs or PLA2 myotoxins
[98].
Resistance to neurotoxic toxins generally involves residue modifications in muscular nicotinic cholinergic receptors (nAChRs) that are targeted by α-neurotoxins. These mechanisms have been primarily reported in front-fanged snakes, transforming colubrids into a novel and outstanding model for exploring toxin resistance and specificity
[99].
Several colubrid species have already been shown to possess venoms with highly specific neurotoxins that exhibit varying potency towards different prey
[29,55]. However, these studies have only been performed on a limited number of genera and the evolutionary consequences of colubrid venom specificity/resistance are yet to be unveiled. Overall, colubrid venoms represent an interesting source to look for novel protein scaffolds and to study evolutionary processes that are poorly understood. An increased amount of venom-related data is needed in order to truly understand the mechanisms that shape venom composition, function, and evolution in this extremely diverse group.
4.2 Colubrid venoms and pharmacological applications
Current research focusing on venoms from rear-fanged snakes is still neglected compared to the amount of work describing the composition and biological diversity of front-fanged snake venoms (Chapters
5 and
10). Therefore, the amount of fully characterized toxins from colubrid venoms is contrastingly low
[11]. The transition from toxin discovery/characterization to the development of a possible therapeutic agent is a very complex process
[88,90]. Historically, venom research has been a great source of proteins and peptides with a vast potential application in numerous biomedical and pharmacological fields (see Chapter 10). Hence, there are several therapeutic compounds in circulation and/or in clinical trials used in humans that have been successfully designed and developed from animal poisons and venoms
[88,90,100–102].
Considering that rear-fanged snake venoms contain a vast number of different proteins and peptides with complex and potent biological activities resembling those found in vipers and elapids, they represent a novel source of potentially bioactive molecules
[28,37,39–41]. The implementation of new extraction methods, the improvement of methodological assays, and the increasing interest in these venoms have resulted in a better understanding of their complex, unique, and new biological activities and molecules
[11,40,103]. The huge taxonomic diversity of colubrids has resulted in a great variety of compositional and functional venom profiles as previously described in this chapter. All of these factors set an enormous potential for the discovery of novel protein families or enhanced biological effects of some compounds in terms of potency or efficacy that may represent an excellent basis for drug design and development
[10,40,103].
Research on rear-fanged snake venoms has revealed a complex variety of biological activities and compositional patterns. However, the most ubiquitous and commonly studied activities in colubrid venoms are proteolytic activity that leads to hemorrhage and coagulation disturbances, and myotoxic and edematogenic activities that resemble the effects observed in
Bothrops venoms
[32–34,42,56] Moreover, some protein families like SVMPs and PLA2 found in colubrid venoms show different toxic and enzymatic potencies compared to the ones found in species of viperids
[32,34]. These functional differences, as well as prey specific toxicities, might be interesting approaches for future research looking for compounds with therapeutic potential
[33,56].
The presence of conserved structures in common toxins (e.g. CRISPs) that are spread throughout many venomous reptiles, together with non-enzymatic proteins known to produce several pharmacological activities, also provides venoms with different and unknown biological properties to be examined
[11,32,37,39]. A prime example of this is
patagonin,a CRISP isolated from the venom of
Philodryas patagoniensis that shows antibacterial activity at very low concentrations without signs of cytotoxic activity
[104,105].
Another very interesting fact that underlines research on colubrid venoms as a great source of potential discoveries is the presence of unique and putative toxins in these venoms
[11,32,33,42]. The presence of these unique protein families provides different research approaches for discovering the biological role and activities from these toxins such as possible new physiological mechanisms, pharmacological targets, and even therapeutic potential.
On the other hand, the diversity of habitats, diets and behaviors found in Colombian colubrids might translate into drastically different toxin scaffolds from that of elapid and viperid venoms. However, the lack of venom-related data has hindered the advance of studies that unveil the pharmacological and functional applications of colubrid venoms. The advance and development of new methods and tools for venom research represents a great opportunity to better understand the biological implications of these toxins and to provide valuable information for possible future novel drug discoveries.
5.
The Colombian perspective
During the last few decades, an interest in research focused on colubrid venoms and envenomation cases has bloomed. The implementation of novel venom extraction methods, including manual and pharmacological protocols, has resulted in greater venom yields. This has eased the development of better transcriptomic and proteomic methodologies that have increased our understanding of the composition and biological properties of colubrid toxins using small quantities of venom
[11,37–40].
Knowledge about these venoms obtained during recent years and the awareness of the potential hazards that some colubrid species represent around the world has led to the implementation of different strategies to record and control envenomation cases including these species
[134,135]. These strategies focus on engaging with communities to increase their awareness, ensuring the availability of efficient and specific medical treatment, strengthening the health system, and establishing interdisciplinary partnerships between the public and private sectors. In many countries, institutions are encouraged to develop security measures to handle accidents involving these species and to achieve a faster and more accurate taxonomical identification in eventual envenomation cases
[135]. In Brazil, for example, colubrid snakebites represent a collective health problem with many cases reported per year and with the awareness that some snakes previously considered harmless could be of medical interest
[35,45,49].
In Colombia, there are around 340 species of snakes and 18% (~20 viperid and ⁓29 elapid) represent species of medical interest
[8,136,137] (See Chapter 1). Colubrids contain the majority of the snake species found in the country (~291 species) and many of these possess an opistoglyphous dentition (enlarged grooved posterior maxillary teeth) connected to the Duvernoy’s venom gland that makes them a potential hazard for humans
[17,36].
However, this group of snakes receives almost no medical or epidemiological interest in Colombia. The lack of ability to identify these species by the community in general and the medical staff, as well as the shortfalls in the surveillance system, are the main constraints facing snakebites caused by non-front-fanged snake species
[10]. An improved surveillance and understanding of these cases would lead to better treatment for the patients and would avoid some practices such as the injection of bothropic or elapidic antivenom that might not be necessary.
Because of minimal medical interest for these species and venoms, there is also a large knowledge gap and a lack of information of the compositional and biological properties of venom samples from Colombian colubrid species
[33,56]. The relevance of the toxinological research of this group lies in the vast diversity of new family proteins/toxins and biological functions that also represent a great fountain of resources for bioprospection and evolutionary/phylogeny approaches
[32,34].
It is necessary to include these species in the snakebite programs in Colombia. They should also be included in epidemiological surveillance strategies. This will lead to the restoration of security measures and protocols for the maintenance of colubrid snakes in captivity and to a better system of notification and registration of accidents that include these species. Along with these data, regional epidemiological studies of these accidents would aid in the identification of the most affected communities and the implementation of educational programs that increase medical awareness of these species. Finally, these approaches will result in increased interest of research groups for investigating these venoms and for carrying out interdisciplinary projects to understand the composition, evolution, and mechanisms of action of these venoms.
Appendix: Material and Methods
The search methods for identifying studies were based on weekly searches from January to June 2024. We performed the searches as follow:
- Weekly searches of google scholar
- Weekly searches of Scielo
- Weekly searches of Scopus
The search strategies contained the following keywords and were combined in different ways using the boolean connectors AND OR: snake venoms, colubrid, Colubridae, Dipsadidae,
Xenodon, Leptophis, Erythrolamprus, Oxybelis, Helicops, Thamnodynastes, Leptodeira, Philodryas, Pseudoboa, Tantilla, Spilotes, Rhinobothryum, Tretanorhinus, Oxyrhopus, Lygophis, transcriptome, proteome, evolution, epidemiology, pharmacology, and drug development.
To compare the relationship between different venom toxin classes in colubrids, we recovered expression data of relevant toxins from published works with colubrid species (both proteomic and transcriptomic)
[29,42,52,54,55,57,132]. We transformed the data into percentages to obtain comparable values for the proportions of each toxin class. Venoms were considered highly neurotoxic or proteolytic if >30% of the identified toxins were linked to those activities. We performed an ancestral state reconstruction analysis using the default parameters of the
fastAnc function implemented in the Phytools package
[138] on the R software using the phylogeny proposed by Zaher et al.
[9] to estimate branch lengths using
Micrurus corallinus and
Bothrops jararaca as ourgroups. Then we built a tree using the
contMap function in Phytools to represent the continuous variation in toxin proportions for each node using a color gradient. Compositional data for
Micrurus and
Bothrops were obtained from previous research
[80,81]. As these data were obtained from various investigations that implemented different methodologies, some lacking proteomic confirmation of reported toxins, our comparisons are merely qualitative and should be interpreted with prudence. A more thoroughly built dataset is needed to draw a robust view of venom variation across colubrids.
Epidemiological data were obtained from manual inspection of National Public Health Surveillance System (Sistema Nacional de Vigilancia en Salud Pública: SIVIGILA, Spanish acronym) reports. Annual cases, clinical manifestations, species identification, and the use of antivenom were checked for all cases. A species was considered to be identified when the report included its full scientific name or a sufficiently specific common name that allowed for it to be classified at the family level. Annual cases were plotted using the PlotR packge developed by Guisande-González
[139], and linear regression models were performed following the procedures described by Guisande-González et al.
[140].