1. Introduction
The family Elapidae is a diverse group of venomous snakes, with nearly 390 species, and most elapids are distributed in the tropics
[1–4]. In the Americas, the family is represented by a monophyletic clade containing the coralsnakes (genera
Micruroides and
Micrurus) and the marine species
Hydrophis
platurus
[1–4]. Coralsnakes comprise almost 90 species and range from the United States to Argentina, but they are absent in Chile and most Antillean islands
[1,2].
The genus
Micruroides is the most northerly distributed coral snake clade, and the only recognized species in this genus,
M. euryxanthus,occurs in Mexico and the United States
[1,2]. The genus
Micrurus is diverse
[1,2]; most species occur in South America, being Brazil and Colombia the countries with the highest species diversity (~ 30 species each)
[1,2,5].
Coralsnakes are widely recognized for their bright coloration and for being highly venomous. Their color patterns consist of rings arranged in three colors, which normally include a light color (white or yellow), a bright color (red or orange), and black; however, some species have atypical color patterns
[5–8]. Four types of ring arrangements are recognized among coral snakes of the genus
Micrurus, and this corresponds to clades in their phylogenetic tree
[2,5,6]. The ring arrangements described below correspond to their body coloration and may or may not occur on the head and tail (Figure 1).
-
Bicolor: rings in the body arranged in two colors, black alternating with a light or bright color. The head and tail in these species normally have black rings arranged between a bright color (red or orange).
-
Monads: rings in the body in three colors, with one black ring between the brightly colored rings, and separated from these by light colored rings. The head and tail in these species normally have rings with a light color and black. The name of this color pattern refers to the number of black rings (one) between the brightly colored ones. This color pattern has evolved independently in
Micruroides and a monophyletic clade of
Micrurus.
-
Triads: rings in the body arranged in three colors, with three black rings, separated by light colored rings, which are together between the bright rings. The head and tail ring arrangement are variable but normally is similar to that of the body. The name of this color pattern refers to the number of black rings (three) between the brightly colored ones. This color pattern occurs independently in one large South American clade and in two Central American species.
-
Incomplete rings: this pattern is almost identical to the bicolored pattern, but features light rings along the body, which shorten down towards the dorsum and normally are not complete; the dorsal coloration is mainly black. This pattern occurs in a group of species known as pygmy coral snakes, which were previously grouped in a different genus,
Leptomicrurus.
.png)
Figure 1. General patterns of the body coloration in Colombian Coral snakes. Bicolor BWB (Micrurus mipartitus): Bicolor pattern with alternating black and white rings. Bicolor RBR (Micrurus multiscutatus): Bicolor with alternating red and black rings. Bicolor B(R/W)B (Micrurus camilae): Bicolor pattern alternating red and black dorsally and alternating white and black ventrolaterally; Monadal RYBYR or RWBWR: Monadal pattern with red rings separated from black rings, alternating with white or yellow rings. Triadial RBWBWBR (Micrurus surinamensis): Triadal with red rings separated by three black rings and two white/yellow rings; Incomplete rings (Micrurus scutiventris): incomplete light rings along the body, which narrow down towards the dorsum and normally are not complete; the dorsal coloration is mainly black. Acronyms: BWB= Black/White/Black. RBR=Red/Black/Red. B(R/W)B= Black/half red and half white/Black. RYBYR= Red/Yellow/Black/Yellow/Red. RWBWR=Red/White/Black/White/Red. RBWBWBR= Red/Black/White/Black/White/Black/Red. RBYBYBR=Red/Black/Yellow/Black/Yellow/Black/Red.
Coralsnake venom is primarily neurotoxic, inducing a neuromuscular blockade that can result in the prey's death through respiratory arrest
[9]. Their venoms exhibit moderate heterogeneity compared to other groups, such as viperids or colubrids, but contain a diverse range of toxin components, with each species having between 4 to 24 different protein families
[2,4,5,9,10]. The main components of these venoms are phospholipases A2 (PLA2), which predominantly mediate presynaptic neurotoxicity, and three finger toxins (3FTx), which are mostly associated with postsynaptic effects
[2]. One of these toxin families is usually dominant over the other in almost all species of coralsnakes, and this creates a phenotypic dichotomy across species
[2,9,11].
The dominance of these two proteins has been suggested to respond to a geographical gradient, with species in the northern range of the genus
Micrurus having PLA2-dominated venoms while those in the southern range having a predominance of 3FTx
[9,11]. This hypothesis has been subject to recent debate, given that some studies have provided contradicting evidence
[2,12]. Likewise, as we will show in this chapter, the geographical pattern of the ratio PLA2/3FTx is blurred when it is confronted to a larger population sampling and increase of species richness. However, an additional evolutionary hypothesis for the observed phenotypic dichotomy has not been proposed.
The phylogenetic relationships of the genus
Micrurus have been discussed in several studies
[3,6,13–16], but advances come at a low pace due to the challenges associated with the high diversity of the group and the cryptic nature of most species
[6,14,15]. In general, most studies have suggested that species with the monadal pattern form a monophyletic group and South American triadal snakes plus the bicolored species also form a clade
[13–15]. The phylogenetic position and monophyly of the pygmy coralsnakes (which have incomplete rings) and the Central American species with triadal patterns (M. elegans) have not yet been elucidated, mainly due to a scarcity of samples and poor genetic resolution
[15,17].
In this chapter, we propose a phylogenetic hypothesis for the coralsnakes of the genus
Micrurus using published sequences and test whether venom composition is associated with phylogenetic and geographic patterns. We also review evidence for and against the longstanding hypothesis that the color patterns of coral snake’s function as warning signals and discuss alternative explanations for the function of coral snake color patterns.
2. Phylogenetic relationships of the New World coralsnakes in the genus
Micrurus
Our maximum likelihood phylogenetic tree recovered New World coralsnakes (Micruroides +
Micrurus) and the genus
Micrurus as monophyletic with high support (Figure 2). Within
Micrurus, two highly supported clades were recovered, one including all the species with monads plus
M. elegans, one of the Central American species with triads; and the second contained all the South American species with triads,
M. mipartitus, the only bicolored species, and
M. narducci, the only species from the old-fashioned genus
Leptomicrurus
[8] (also known as pygmy coralsnakes, which are herein classified as coralsnakes with a color pattern of incomplete rings).
Three main subclades can be recognized within the clade containing the triadal species: one highly supported subclade containing species in the
M. lemniscatus complex (M. diutius,
M. carvalhoi,
M. filiformis,
M. helleri, and
M. lemniscatus),
M. surinamensis, and
M. ortonii (clade T1, Figure 2), which is sister to
M. narduccii, forming a poorly supported clade. A second highly supported subclade that includes
M. dissoleucus and
M. mipartitus (clade T2, Figure 2). A third weakly supported clade that includes species of the
M. frontalis group (M. altirostris,
M. baliocoryphus,
M. brasiliensis,
M. frontalis,
M. ibiboboca,
M. cf.
ibiboboca, and
M. pyrrhocryptus), plus
M. decoratus and
M. obscurus (clade T3, Figure 2).
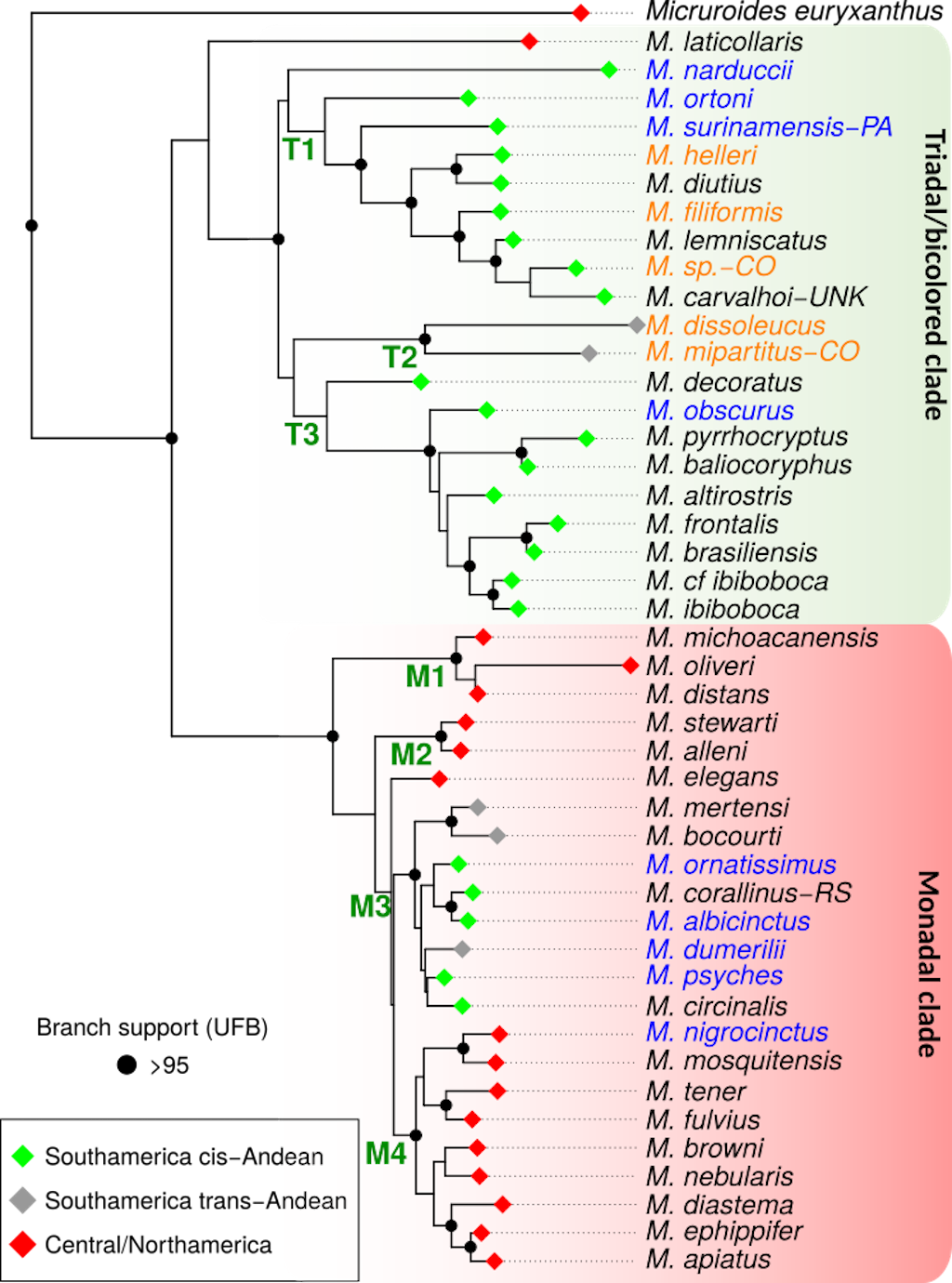
Figure 2. Maximum likelihood tree obtained via IQ-Tree, based on the concatenated alignment of four mitochondrial (12S, 16S, cytb and ND4) and one nuclear (Cmos) gene (3,175 bp). The colors of the node circles indicate ultrafast bootstrap support values, red: >95, black: <95. Color at tips indicates species that occur in Colombia; orange names indicate sequences from Colombian specimens, and blue names indicate sequences from non-Colombian samples. Notations for some samples (CO, PA, RS, etc) are specified in the appendix.
A total of four main subclades were recovered with high support for the clade containing the monadal species: the first contained taxa in the
M. distans group (M. distans,
M. michoacanensis, and
M. oliveri; clade M1, Figure 2); the second contained
M. stewarti and
M. alleni (clade M2), two central American species; the third contained all the South American monadal species (M. albicinctus,
M. bocourti,
M. circinalis,
M. corallinus,
M. dumerilii,
M. mertensi,
M. ornatissimus, and
M. psyches; clade M3, Figure 2); and the fourth contained the remaining Central and North American species (clade M4, Figure 2). In our phylogenetic tree, 14 of the 46 species sampled occur in Colombia, and six of these sequences were derived from Colombian specimens (Figure 2).
In general, our phylogenetic hypothesis is in line with most recent revisions, finding as monophyletic the monads clade with the bicolored and the triadal clades forming a monophyletic clade
[14-17,19–23]. The relationships of
< a target="_blank" href="/en/Paginas/files/Micrurus-narduccii.aspx"> Micrurus narducii, the only representative of the old-fashioned genus
Leptomicrurus, remain ambiguous among studies, while in our tree it falls within the triadal clade, as in Jowers et al.
[14], other recent studies found it as sister to the triadal clade
[20,21,23].
The phylogenetic hypothesis presented herein (Figure 2), represents the largest phylogenetic proposal for the new world coralsnakes in the genus
Micrurus, including 44 (plus two unclassified taxa,
M. sp., and
M. cf.
ibiboboca), out of the 83 currently recognized species on the genus (53%)
[1]. With this sampling and the phylogenetic context some biogeographic patterns arise.
The clade grouping the triadal + bicolored + incomplete rings is composed almost exclusively of South American species (Figure 2), except for the basal species,
M. laticollaris, which is distributed in Mexico
[24,25]. Nevertheless, the position of
M. laticollaris is poorly supported and its position might change with more evidence. Within the South American cluster, all species are strictly distributed in the cis-Andean region (i.e. Orinoquia ecoregion and Amazon ecoregion, Figure 3), except for two taxa which form a highly supported clade,
M. dissoleucus and
M. mipartitus, both distributed in the trans-Andean region (i.e. across or west of the Andes, Figure 3)
[24,25]. This suggests that this clade diversified in the cis-Andean region and most likely colonized the trans-Andean region a single time.
On the other hand, the bicolored clade groups taxa from the whole American continent, but with most taxa distributed in Central America (Figure 2). Within this clade, there is a lineage that groups all South American species, which might be suggestive of a single colonization event to South America for this clade (Figure 2). Nevertheless, some other species in the monadal clade, also enter northern South America, like
M. nigrocinctus and
M. clarki
[24,25]. Despite the internal relationships that are not well supported within the South American monadal lineage, the retrieved topology suggests at least two crosses of the Andes (e.g. from the trans to the cis-Andean region, or vice versa, Figure 2).
Even though it is widely known in the literature that the monadal clade is mainly comprised of Central American species, and the triadal clade is composed mainly of South American/cis-Andean taxa
[24,25], no previous study has shown or discussed the biogeographic patterns of diversification within
Micrurus, based on a phylogenetic framework. Also, the fact that the main clades have most of their diversity grouped in specific regions (e.g. Central- or South America), might have led to some authors (i.e. Lomonte et al.,
[9]) to suggest latitudinal variation in the venom composition within
Micrurus, which is further discussed in the next section.
3. Compositional diversity across Micrurus venoms
We compiled literature data of 34 proteomes belonging to 28
Micrurus species (Table 1).
Micrurus corallinus,
M. mipartitus,
M. helleri, and
M. surinamensis had two published proteomes, and
M. carvalhoi had three published proteomes. Only six of the available proteomes came for Colombian specimens (M. dumerilii,
M. mipartitus-CO,
M. sangilensis,
M. helleri-CO,
M. medemi, and
M. sp.-CO).
Based on the 34 proteomes, a total of 24 protein families was reported among
Micrurus venoms, and PLA2 and 3FTx were the dominant venom components in all proteomes, as has been previously found in multiple studies
[26]. Fifteen toxin-like protein classes were identified in the proteomes of
M. carvalhoi-UNK and
M. pyrrhocryptus, which was the highest among all proteomes in the triadal clade. Twelve toxins were identified in the proteomes of
M. corallinus-RJ and
M. paraensis, which were the most diverse among all proteomes in the monadal clade (Table 1). In general, PLA2-dominated venoms were more common in the monadal clade, whereas 3FTx-dominated venoms were more common within the triadal clade (Figures 3 and 4).
Our comparisons of available compositional data for
Micrurus species revealed intraspecific variation in both the occurrence and relative abundance of toxins in
M. carvalhoi, M. corallinus,
M. helleri,
M. mipartitus, and
M. surinamensis, which had available data for multiple populations or individuals (Table 1). In these species, we noted that the dominant toxin, either 3FTx or PLA2, varied between individuals across their geographic distribution, without any clear pattern (Figure 2). Differences in the relative abundance of 3FTx and PLA2 (|%3FTx-%PLA2|) were more pronounced in the triadal + bicolored clade (~44%), than in the monadal clade (~29%; Table 1). This indicates that proteomes from the triadal + bicolored clade tend to exhibit more extreme phenotypes, favoring a marked dominance of either PLA2 or 3FTx, whereas the monadal groups possess more balanced PLA2 or 3FTx proportions. Moreover, the genus
Micruroides shows a higher proportion of 3FTx, suggesting that the emergence of PLA2-dominated venoms could be a derived phenotype within coralsnakes.
Table 1. Venom proteome composition from available proteomes in the literature. Values are shown in percentages of total spectra to allow for compositional comparisons between species. Red and orange cells indicate higher proportions of each toxin, and yellowish and green cells indicate lower proportions of each toxin. Species are catalogued by their clade (monadal, triadal/bicolored, or the genus
Micruroides). Only major toxins are shown. SVMP: Snake venom metalloproteinases, LAO: L-amino acid oxidases, Kun: Kunitz inhibitors, CTL: C-type lectins, SP: Serineproteases, Oha: Ohanin.
| Species |
Clade |
PLA2/3FTx delta |
3FTx |
PLA2 |
SVMP |
LAO |
Kun |
CTL |
SP |
Oha |
Minor toxins |
| Micruroides euryxanthus |
Micruroides |
48.1 |
62.3 |
14.2 |
|
|
|
|
|
|
0 |
| Micrurus alleni |
Monadal |
66.4 |
77.3 |
10.9 |
1.2 |
3 |
|
1.8 |
|
0.2 |
1 |
| Micrurus browni |
Monadal |
12.2 |
50.4 |
38.2 |
1.4 |
0.4 |
4.2 |
1.1 |
0.3 |
0.2 |
2.9 |
|
Micrurus clarki |
Monadal |
11.7 |
48.2 |
36.5 |
1.6 |
3.8 |
0.9 |
1.1 |
1 |
|
0.4 |
| Micrurus corallinus-RJ |
Monadal |
3.71 |
34.83 |
38.54 |
10.38 |
5.42 |
0.46 |
|
|
|
10.37 |
| Micrurus corallinus-UNK |
Monadal |
69.8 |
81.7 |
11.9 |
2.4 |
2.3 |
|
0.9 |
0.8 |
|
0 |
|
Micrurus dumerilii |
Monadal |
23.9 |
28.1 |
52 |
1.8 |
3.1 |
9 |
1.1 |
1.9 |
|
0.4 |
| Micrurus fulvius |
Monadal |
33.4 |
24.9 |
58.3 |
|
|
|
|
|
|
0 |
| Micrurus mosquitensis |
Monadal |
33.1 |
22.5 |
55.6 |
2.6 |
2.8 |
9.8 |
1.3 |
0.5 |
|
1 |
|
Micrurus nigrocinctus |
Monadal |
10 |
38 |
48 |
4.3 |
2.3 |
|
2.2 |
0.7 |
3.8 |
0.5 |
| Micrurus paraensis |
Monadal |
53.58 |
12.31 |
65.89 |
7.97 |
3.13 |
3.43 |
4.95 |
|
|
2.3 |
|
Micrurus sangilensis |
Monadal |
12.71 |
17.69 |
30.4 |
11.81 |
9.17 |
2.97 |
6.27 |
5.79 |
2.27 |
13.63 |
| Micrurus tener |
Monadal |
8.1 |
37.5 |
45.6 |
|
|
|
|
|
|
0 |
| Micrurus altirostris |
Triadal + Bicolored |
65.8 |
79.5 |
13.7 |
0.9 |
1.2 |
2.1 |
0.1 |
|
|
2.5 |
| Micrurus carvalhoi-GO |
Triadal + Bicolored |
46.31 |
2.25 |
48.56 |
6.29 |
2.01 |
12.61 |
2.64 |
2.12 |
21.51 |
1.99 |
| Micrurus carvalhoi-SP-B |
Triadal + Bicolored |
49.2 |
71.3 |
22.1 |
0.8 |
1.5 |
0.03 |
|
|
|
0.01 |
| Micrurus carvalhoi-SP-J |
Triadal + Bicolored |
56.5 |
76.7 |
20.2 |
1.13 |
1.3 |
0.03 |
|
|
|
0.175 |
| Micrurus frontalis-BR-SP |
Triadal + Bicolored |
6.8 |
42.4 |
49.2 |
0.96 |
0.4 |
1.3 |
0.06 |
|
|
|
|
Micrurus helleri-CO |
Triadal + Bicolored |
26.53 |
14.1 |
40.63 |
13.1 |
8.64 |
2.2 |
0.58 |
5.24 |
|
15.52 |
|
Micrurus helleri-EC |
Triadal + Bicolored |
54.3 |
17.8 |
72.1 |
|
5.1 |
0.5 |
|
|
|
4.5 |
| Micrurus ibiboboca |
Triadal + Bicolored |
53 |
13.5 |
66.5 |
2.5 |
2.8 |
9.4 |
0.8 |
|
|
3.4 |
|
Micrurus lemniscatus |
Triadal+Bicolored |
14.95 |
34.33 |
19.38 |
4.93 |
3.48 |
11.69 |
2.59 |
11.43 |
9.32 |
2.86 |
|
Micrurus medemi |
Triadal + Bicolored |
25.45 |
17.69 |
43.14 |
9.63 |
7.48 |
5.35 |
7.83 |
0.39 |
|
8.5 |
|
Micrurus mipartitus-CO |
Triadal + Bicolored |
32.1 |
61.1 |
29 |
1.6 |
4 |
1.9 |
1.1 |
1.3 |
|
0 |
|
Micrurus mipartitus-EC |
Triadal + Bicolored |
44.3 |
63.4 |
19.1 |
6 |
8.4 |
|
|
|
|
3.1 |
| Micrurus multifasciatus |
Triadal + Bicolored |
74.8 |
83 |
8.2 |
3.6 |
3.2 |
1.9 |
|
|
|
0.3 |
| Micrurus pyrrhocryptus |
Triadal + Bicolored |
10 |
27 |
17 |
12 |
5 |
5 |
7 |
7 |
2 |
10 |
| Micrurus ruatanus |
Triadal + Bicolored |
16.6 |
46.4 |
29.8 |
2.7 |
3.4 |
10.6 |
2 |
1.2 |
0.4 |
0.5 |
| Micrurus sp-AL |
Triadal + Bicolored |
31.7 |
26.1 |
57.8 |
1.8 |
2 |
5.6 |
2 |
|
0.38 |
3.0 |
| Micrurus sp-CO |
Triadal + Bicolored |
41.4 |
21.1 |
62.5 |
|
|
5.9 |
0.5 |
|
|
0 |
| Micrurus spixii |
Triadal + Bicolored |
54.56 |
9.48 |
64.04 |
6.57 |
4.78 |
0.74 |
7.37 |
|
3.73 |
3.27 |
| Micrurus spixii-BR-PA |
Triadal + Bicolored |
19.1 |
56.5 |
37.4 |
|
5.1 |
1 |
|
|
|
|
| Micrurus sp-RJ |
Triadal + Bicolored |
94.1 |
94.1 |
0 |
|
0.07 |
0.98 |
|
|
|
0.97 |
|
Micrurus surinamensis-BR-RO |
Triadal + Bicolored |
91.2 |
95.4 |
4.2 |
|
0.4 |
|
|
|
|
|
|
Micrurussurinamensis-PA |
Triadal + Bicolored |
83.54 |
90.1 |
6.56 |
0.11 |
0.19 |
2.55 |
0.32 |
|
|
0.19 |
|
Micrurussurinamensis-TO |
Triadal + Bicolored |
3.86 |
36.84 |
32.98 |
1.23 |
0.32 |
25.26 |
1.66 |
|
|
1.7 |
| Micrurus tschudii |
Triadal + Bicolored |
91.1 |
95.2 |
4.1 |
|
0.7 |
1.6 |
|
|
|
0 |
AL (Alagoas, Brazil), B (Baurú, Brazil), BR (Brazil), CO (Colombia), EC (Ecuador), GO (Goiás, Brazil), J (Jaú, Brazil), PA (Pará, Brazil), RJ (Rio de Janeiro, Brazil), RO (Roraima, Brazil), SP (São Paulo, Brazil), TO (Tocantins, Brazil), UNK (Unknown exact location).
M. sp.-CO: A specimen of
Micrurus aff. lemniscatus, but with a significant genetic distance from its congeners; see Hurtado-Gómez
[13] for details.
Several studies have suggested a compositional dichotomy in the venom of
Micrurus, in which the dominant toxin (either PLA2 or 3FTx) alternates between species across a geographic spectrum
[9,11]. However, such a dichotomy is not evident when we map the variation of venom composition across the geographic range of
Micrurus (Figure 3).
Latitudinal geographic location alone thus appears to be a poor predictor of a predominance of 3FTx or PLA2 in their venom proteomes. However, some species have consistently higher levels of 3FTx compared with PLA2 and vice versa, and this might be explained by their phylogenetic relationships or ecological convergences. To assess whether the predominance of 3FTx/PLA2 is determined by phylogenetic signal, we mapped the proportions of each toxin in the venom proteome to the species phylogeny (Figure 4). As a result, we did not observe any clear pattern of 3FTx/PLA2 predominance across the phylogeny of coralsnakes. Both the triadal and the monadal clades have species with venom proteomes dominated by either 3FTx or PLA2. However, most species possess a balanced ratio between both toxins, with medium proportions of each one. This balanced phenotype is the most common in both clades, and extreme phenotypes, in which one toxin dominates, have emerged in specific groups within both clades, regardless their geographical origin.
.png)
Figure 3. Geographic distribution of
Micrurus samples with available venom proteomes. 3FTx and PLA2 proportions are shown for each sample. Color patterns are indicated by colored points above the geographic location of each sample. AL (Alagoas, Brazil), B (Baurú, Brazil), BR (Brazil), CO (Colombia), EC (Ecuador), GO (Goiás, Brazil), J (Jaú, Brazil), PA (Pará, Brazil), RJ (Rio de Janeiro, Brazil), RO (Roraima, Brazil), SP (São Paulo, Brazil), TO (Tocantins, Brazil), UNK (Unknown exact location).
M. sp.-CO: A specimen of
Micrurus aff. lemniscatus, but with a significant genetic distance from its congeners; see Hurtado-Gómez
[13] for details.
This might indicate that ecological factors, perhaps related to diet, microhabitat use, abiotic features, climate, or other environmental variables could be determining the dominant toxins in these snakes. Additional ecological studies could help clarify the evolutionary origins of the observed variation in the venoms of
Micrurus. Moreover, a recent work reported elevated proportions of Snake Venom Metalloproteinases (SVMPs) in two Colombian species of the genus
[27]. These toxins are commonly associated with viper venoms and are linked with local effects (e.g., bleeding, edema, necrosis) not normally observed with coral snake envenomations
[2,10,28]. These findings are particularly relevant to the diagnosis of coral snake envenoming, given that bites from these snakes with profusive bleeding or local manifestations might be misclassified as other type of envenomation (i.e. bothropic
see Chapter 9), which can lead to serious clinical consequences.
In summary, available proteomic information for
Micrurus venoms reveals that our knowledge of the factors driving their compositional variance remains scarce and much compositional variation is likely yet to be discovered (e.g., some coralsnakes might have higher than expected proportions of SVMPs in their venoms). Studies with increased sampling of coralsnakes that occupy diverse environments are needed to improve our understanding of the compositional range of coral snake venoms and their local medical implications (see Chapters
5,
6, and ).
.png)
Figure 4. The phylogenetic distribution of the proportions of 3FTx/PLA2 in the venom proteomes of coralsnakes. Terminal branches with names in bold font correspond to species with published proteomic data. All others were estimated using the anc.ML function of the phytools package in R (see Appendix). Blue branches indicate higher proportions of each of the toxins (PLA2 on the left and 3FTx on the right). AL (Alagoas, Brazil), B (Baurú, Brazil), BR (Brazil), CO (Colombia), EC (Ecuador), GO (Goiás, Brazil), J (Jaú, Brazil), PA (Pará, Brazil), RJ (Rio de Janeiro, Brazil), RO (Roraima, Brazil), SP (São Paulo, Brazil), TO (Tocantins, Brazil), UNK (Unknown exact location). M. sp.-CO: A specimen of
Micrurus aff. lemniscatus but with a significant genetic distance from its congeners see Hurtado-Gómez
[13].
4. Aposematism and coralsnake mimicry
Coralsnakes are often brightly colored and seem to defy expectations that camouflage is the main driver of the coloration of non-social animals. The function (i.e., the effect that selected for the evolution and maintenance of a trait) of these unusual colors has been the subject of scientific inquiry for over 150 years
[18,30]. The function of the color patterns of coralsnakes has long been assumed to be aposematic, meaning that their color patterns are thought to serve as warning signals that advertise their danger to predators
[43,44]. False coralsnakes have also been assumed to have similar color patterns because a resemblance to highly venomous coralsnakes is thought to protect them from predators, which confuse false coralsnakes with more dangerous coralsnakes
[35,45,50]. These assumptions are widespread among biologists, especially those engaged in evolutionary ecology studies of coralsnakes and false coralsnakes
[29].
A recent meta-analysis has shown that aposematism and mimicry in coralsnakes are supported by most studies since the 1980s despite gaps in evidence
[30], and a recent meticulous analysis of aposematism and mimicry in coralsnakes and false coralsnakes has shown that the theoretical and empirical basis of these ideas has serious problems
[31], suggesting that these longstanding assumptions merit reconsideration.
In light of this lack of consensus, we provide a field herpetologist perspective on this controversy. We highlight some of the main unresolved problems and possible alternative explanations for the functions of coral snake color patterns. In contrast to a recent review by Almeida et al.
[29], we take a more critical perspective on past work and concur with Mouy
[31] that there is a need for increased focus on alternative explanations for the functions of coral snake color patterns.
4.1 Summary of the evidence for aposematism and coral snake mimicry
Observations of avoidance of coralsnake color patterns by predators are considered by proponents of aposematism and coral snake mimicry to provide some of the main evidence
[29,30]. Predator avoidance has been inferred experimentally using field experiments with artificial prey. In these experiments, replicas with coral snake color patterns and non-coral snake color patterns are placed in natural habitats, where they are exposed to free-ranging predators
[32-42]. After a certain period of time, the replicas are scored as attacked or not based on the presence of tooth or beak marks, and these data are then used to infer the relative attack rates of the predator community on coral snake color patterns and thus the degree to which these patterns are protected from predation
[32-42].
These experiments have been conducted in various regions of the Americas and have often shown that the frequency of tooth and beak marks on replicas with coralsnake color patterns is much lower than that on replicas with non-coralsnake color patterns, and this has been interpreted as evidence that coralsnake color patterns provide protection from predation by avian and mammalian predators
[32-38]. Studies of coralsnake color patterns using artificial prey have not yet been conducted in Colombia.
Avoidance of coralsnake color patterns has also been inferred through laboratory experiments. Young turquoise-browed motmots (Eumomota superciliosa) and great kiskadees (Pitangus sulphuratus) from Costa Rica raised in captivity were presented with dowels painted with coralsnake color patterns, and they were reported to display strong negative reactions to dowels painted with coralsnake color patterns
[43,44]. However, these negative reactions were not observed when these birds were presented with dowels painted with single colors, rings with non-coralsnake colors, and stripes with coral snake colors. As these birds had no prior exposure to coral snake color patterns, the aversive reactions observed have been widely cited as evidence of innate avoidance of coralsnake color patterns
[45,46].
Correlations in color patterns between coralsnakes and false coralsnakes have also been suggested to provide support for aposematism and mimicry hypotheses. The color patterns of various coralsnake and false coralsnake species have been shown to vary geographically in parallel with each other. For example,
Erythrolamprus guentheri in southeastern Ecuador and Peru have color patterns that resemble
Micrurus langsdorffi and
Micrurus steindachneri, which have monadal patterns with red and black rings of the same width separated by white rings
[45]. However, at localities in Peru where it co-occurs with the bicolor
Micrurus margaritiferus, which have bicolor patterns with speckled white rings, some individuals of
E. guentheri bear this same pattern
[45].
Similarities in the color pattern of
Oxyrhopus vanidicus with local coralsnakes (i.e.,
Micrurus hemprichii) have been observed in Colombia on the eastern slopes of the Cordillera Oriental as well as on high savannah plateaus of the Orinoquia ecoregion (pers. obs. J.D. Lynch and Teddy Angarita-Sierra). No studies examining correlations in color patterns between false coralsnakes and coralsnakes have been conducted in Colombia; the high diversity in shared color patterns among several coralsnakes and false coralsnakes in the cis-Andean and trans-Andean regions suggests that their color patterns might be correlated in Colombia (Table 2). The study of Bosque et al
[47] showed that the color patterns and geographic distributions of red and black banded
Oxyrhopus species and coralsnakesin South America is associated with the presence and richness of
Micrurus species with the same color patterns
[47].
Variation in the color patterns of
Atractus latifrons and
Erythrolamprus aesculapii has been shown to be associated with the distributions of local
Micrurus species
[48], and the color patterns of various triadal
Micrurus species in Brazil have been shown to be correlated
[49]. Another study integrating distribution and phylogenetic data showed that the evolution of coral snake color patterns among all New World coralsnakes and false coralsnakes is correlated in space and time; specifically, the evolution of coral snake color patterns in false coralsnakes only occurred after the arrival of coralsnakes to the New World
[50].
Table 2. Coralsnakes of Colombia with cis-Andean and trans-Andean distributions and their co-occurring false coralsnakes sharing similar patterns. Color pattern types (following scheme adapted from Mouy
[31]) are indicated in parentheses.
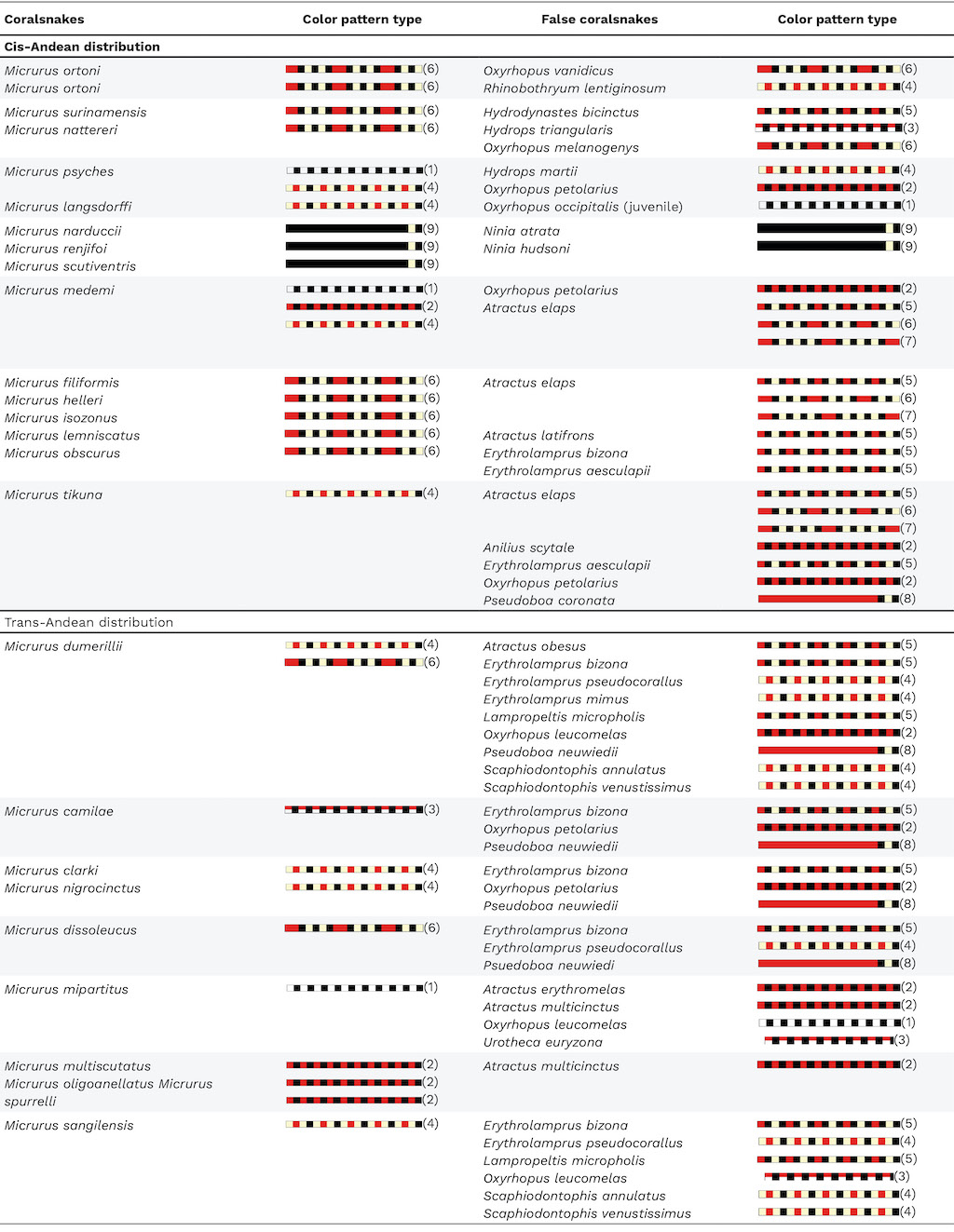
Pattern 1: Bicolor with alternating black and white rings; Pattern 2: Bicolor with alternating red and black rings; Pattern 3: Alternating red and black dorsally and alternating white and black ventrolaterally; Pattern 4: Monadal with red rings separated from black rings; Pattern 5: Monadal with red rings separated from white/yellow rings; Pattern 6: Triadal with red rings separated by three black rings and two white/yellow rings; Pattern 7: Tetradal with red rings separated by four black rings and three white/yellow rings; Pattern 8: Red body with white/yellow nuchal collar between two black rings; Pattern 9: Black body with white/yellow nuchal collar.
4.2 Outstanding issues
A robust, unified framework with clear criteria for determining whether coral snake color patterns have aposematic and mimetic functions is lacking. Consequently, aposematic and mimetic functions of coral snake patterns are assumed a priori in many studies. Although the aforementioned support for aposematism and coralsnake mimicry comprises evidence that is consistent with aposematic and mimetic functions, this evidence is not a perfect match with theory.
What we consider to be the main problems with the aposematism and mimicry hypotheses in coralsnakes and false coralsnakes are the inconsistencies between the putative characteristics of prototypical aposematic and mimetic species
[46] and the natural history and behavior of these snakes
[24]. For aposematism and mimicry to be effective defenses in coralsnakes and false coralsnakes, the color patterns of coralsnakes must be perceived by their avian and mammalian predators (i.e., predators can actually see the colors) and they should be well defended (i.e., coralsnakes are not profitable prey). In the case of learned avoidance, interactions of these predators with coralsnakes should be sufficiently frequent for predators to learn that coralsnakes are unprofitable and for the learning to be reinforced
[46], which means that coralsnakes should occur in microhabitats/time periods in which they would be frequently exposed to their predators to advertise their danger. If these assumptions are not met, aposematism as an explanation for warning coloration becomes untenable. Our own observations suggest that these assumptions are not all met for coralsnakes.
We find that coralsnakes are not frequently active or not readily observable when active on the surface and seem to spend much of their time in microhabitats that are inaccessible to mammalian and avian predators (e.g., under fallen objects or underground). We also find that most coralsnakes are highly inoffensive and reluctant to bite, and this is consistent with observations made by other coral snake researchers
[6,11,51]. This might contribute, at least in part, to explaining the very low incidence of snakebite accidents caused by coralsnakes in Colombia (<1%) and several other countries in the Americas (see Chapter 9). Although some coralsnakes are active by day, many are active at night or twilight conditions when the perception of their warning color patterns by avian predators would be compromised
[6]. Most coralsnakes attempt to flee after we first find them, and many engage in erratic movements after being handled; such behaviors would not be expected for well-defended, aposematic species.
Another serious problem with aposematism and mimicry as explanations of coralsnake color patterns is the frequent occurrence of false coralsnakes in allopatry with coralsnakes, including some “mimic” species that occur far outside the range of any coral snake
[52]. Several explanations have been proposed to explain the presence of allopatric mimics
[52]. These basically either propose that allopatric mimics are not really mimics (e.g., the functions of the color patterns of allopatric mimics differ in allopatry and sympatry with coralsnakes) or that mimicry can occur but via highly unreasonable means (e.g., generalized avoidance).
We have observed no differences in the defensive behavior nor activity patterns of false coralsnakes in sympatry and allopatry with coralsnakes that would suggest any difference in the defensive function of their color patterns. We are also not aware of any studies that have demonstrated a generalized avoidance of coral snake color patterns by any predator, much less how such an avoidance could be genetically programmed (see discussion in Mouy
[31]). The existence of allopatric coralsnakes and false coralsnakes with the same color patterns is also difficult to explain by aposematism and mimicry. Perhaps the most striking case is that of
Micrurus camilae (endemic to Colombia),
Scolecophis atrocinctus (Central America: Guatemala, Honduras, El Salvador, Nicaragua, Costa Rica),
Siphlophis cervinus (trans and cis-Andean regions of tropical South America),
Lygophis elegantissimus (endemic to Argentina), and
Sonora semiannulata (United States and Mexico)
[31].

Figure 5. Geographically distant coralsnakes and false coralsnakes with a similar color pattern (pattern 3 in Figure 1). (A)
Micrurus camilae, photo by José Viera | Ex-situ project. (B)
Scolecophis atrocinctus, photo by Antonia Pachmann. (C)
Sonora semiannulata photo by Ben Thesing. (D)
Siphlophis cervinus, photo by William W. Lamar.
The extensive variation and polymorphism in coralsnake color patterns are difficult to explain under aposematism and mimicry, as diversity in color patterns would only increase the difficulty for predators to learn or acquire aversion of any particular color pattern. This, coupled with the low frequency with which coralsnakes and false coralsnakes are likely encountered by avian and mammalian predators, indicates that the color pattern diversity has an alternative explanation.
The evolution of highly complex color patterns is also problematic to explain via aposematism and mimicry, which would seem to favor the evolution of simple color patterns that would facilitate learning or the evolution of innate recognition. Curiously, some of the most conspicuously colored coralsnakes (e.g.,
Micrurus distans,
Micrurus averyi) and false coralsnakes (e.g.,
Drepanoides anomalus,
Oxyrhopus occipitalis) with perhaps the simplest and easily learnable signals are chiefly nocturnal when their signals would be least visible to mammalian and avian predators.
Several studies have not found any evidence of avoidance of coralsnake color patterns compared with non-coralsnake color patterns. For example, some artificial prey studies employing plasticine replicas have found no evidence a significant reduction in the number of tooth and beak marks on replicas with coral snake color patterns compared with replicas without coral snake color patterns
[39-42]. Coatis (Nasua narica) exposed to live
Micrurus nigrocinctus in a laboratory experiment exhibited no negative responses or aversion to coral snake color patterns
[53]. A recent study in which plasticine models of various snake patterns, including coral snake color patterns, were monitored with camera traps showed that most of the predators that left marks on the replicas were not vertebrate predators; furthermore, the few vertebrate predators that attacked the replicas did not behave in a manner consistent with observations of interactions with natural prey, which suggests that the conclusions of previous studies employing artificial prey techniques should be called into question
[54].
Although coralsnakes have been documented to be preyed upon by various mammalian and avian predators
[55-58], none of these documented predators would likely be strong agents of selection for coralsnake color patterns. For example, some predators, such as
Herpetotheres cachinnans (laughing falcon), can consume any snake, even more well-defended ones such as
Bothrops and
Crotalus species
[59]. Other predators that are not considered snake specialists, such as
Buteo jamaicensis (red-tailed hawk), are also capable of consuming well-defended snakes, such as rattlesnakes of the genus
Crotalus
[60]. The overall rarity of observations of predation of coralsnakes by their widely assumed selective agents (mammals and birds) for aposematism and mimicry over the last 100 years further suggests that they are not major selective agents for coral snake color patterns. Rather than dismissing these findings and observations as innocent exceptions, the possibility that herpetologists have long been mistaken about the function of coral snake color patterns merits serious consideration.
4.3 Aposematism and coral snake mimicry alternatives
Several alternative hypotheses to aposematism and coral snake mimicry have been proposed, but most have not been widely cited nor critically examined. One proposal is that the color patterns of coralsnakes mimic other organisms, such as millipedes, or that coralsnakes are mimicking mildly venomous false coralsnakes, such as
Erythrolamprus species
[45,61]. A mimetic relationship between snakes and millipedes seems unlikely given that most vertebrate predators could probably easily distinguish between them. Moreover, a resemblance between millipedes and snakes with coral snake color patterns is not a widespread phenomenon in the Americas. Coralsnakes are also unlikely mimics of mildly venomous snakes given that coralsnakes can be equally “mild” to predators in the sense that they are incapable of reliably delivering a deadly bite (See Chapter 5). Coralsnakes also occur in many areas without mildly venomous “models.”
The flicker-fusion hypothesis proposes that the rings of snakes’ aid escape from predators by forming a motion blur when the rate of transition of their rings exceeds the flicker-fusion rate of predator eyes; this can cause snakes to appear stationary while moving, which can reduce the success of capture of snake prey by predators
[62]. Although the flicker-fusion hypothesis sounds plausible in that it could potentially provide protection against certain predators, it provides no explanation for the complexity and bright colors of snakes with coral snake color patterns.
Another alternative is that the color patterns serve a direct protective function
[63]. How these color patterns might function in this manner against mammalian and avian predators remains unclear. Mouy
[64] proposed a novel hypothesis for explaining the evolution of coral snake color patterns, the “ophiophagy hypothesis,” which proposes that the rings and colors of coralsnakes and false coralsnakes serve a direct protective function against snake predators. In support of this idea, Mouy
[64] suggested that predatory pressure on snakes in the Americas by ophiophagous snakes might be disproportionately high compared with ophiophagous non-snake predators (i.e., birds and mammals). He then conducted an analysis of snakes with specialized diets of elongated vertebrates and snakes with red and rings and found that there was an association between ophiophagy, red coloration, and ringed patterns, suggesting that the evolution of coralsnake color patterns as a defense against ophiophagy is plausible. Another recent study using a different analytical approach also demonstrated an association between foraging strategy and contrasting colorations in elapid and dipsadine snakes
[65], which provides additional support for the ophiophagy hypothesis.
Mouy
[66] also described a possible mechanism for how coral snake color patterns could thwart predation by ophiophagous snakes. He proposed that rings interfere with the focusing ability of snakes, which have rigid ball lenses, and used a series of simulations to show that spherical aberrations generated by ball lenses can produce local maxima in contrast that would impede the lens from bringing the rings into focus. He further proposed that the function of red in coral snake color patterns is to restrict the vision of snakes in low light environments to rods (photoreceptor cells), which provide a less temporally and spatially accurate vision.
If these functions of rings and red are assumed, the diversity and complexity of coral snake color patterns becomes relatively straightforward to explain. Snakes with lots of red (e.g.,
Pseudoboa neuwiedii, juvenile
Clelia clelia, Drepanoides anomalus, and
Oxyrhopus occipitalis) would likely be preyed upon by nocturnal ophiophagous snakes that rely heavily on rod vision. This is consistent with our observations that these snakes are exclusively nocturnal and relatively fast-moving, which would be particularly important for evading predators with a slow rod-based vision vulnerable to saturation (i.e., the failure of rods to respond beyond certain levels of illumination).
These snakes also appear to avoid moving in open areas under moonlight, which makes sense given that greater light levels could potentially reduce the efficacy of red as a defense. By contrast, snakes with lots of rings (e.g., triadal snakes) are likely preyed upon by ophiophagous snakes with various types of vision, as their patterns would provide protection both through the effect of rings and red. Consistent with this, many of the triadal snakes (e.g.,
Micrurus elegans) have more variable activity patterns and have less red at high elevations (>1000 m above sea level) when temperatures are only conducive to diurnal activity (C.K. Akcali pers. obs.).
To provide preliminary insights into whether coralsnakes would be vulnerable to spherical aberrations, we scanned the eye of a coral snake specimen (Micrurus mipartitus) using an optical coherence tomography system (Figure 6).
%20(1).png)
Figure 6. Averaged optical coherence tomography B-scan of the left eye of
Micrurus mipartitus (Panama, Bocas del Toro, Isla San Cristobal; USNM 348250) along the dorsal–ventral axis. sl: supralabial scale; so: supraocular scale; sp: spectacle; co: cornea; ir: iris; le: lens re: retina, vi: vitreous. Horizontal spectacle diameter: 1.38 mm; Equatorial lens diameter: 0.765 mm.
The scan revealed thatthe anterior surface of the lens of
M. mipartitus was subspherical as has been observed in other snakes
[67], which indicates that the eyes of
M. mipartitus would in fact be vulnerable to experiencing spherical aberrations. Detailed studies of the eye morphology and visual systems of coralsnakes and false coralsnakes are needed to determine the extent to which coral snake color patterns could prove effective as direct defenses against snake predators.
Although much work is needed to evaluate the ophiophagous hypothesis and possible mechanisms for how coral snake color patterns could provide a protective function, the explanatory power of the framework proposed by Mouy
[64,66] is immensely greater than that of all other explanations for the evolution of coralsnake color patterns proposed to date, including aposematism and mimicry. It provides compelling explanations for the diversity and complexity of coral snake color patterns and is free of many of the traditional problems of aposematism and mimicry.
It can also explain various curiosities that alternatives provide no explanation for, including: (1) why certain color patterns, such as red and white rings without any black rings, have never evolved (the proposed function requires contrasting rings, and red and white rings might not provide sufficient contrast under most lighting conditions); (2) why white/yellow rings are rarely more than 50% of the body unlike red (the exclusive purpose of white/yellow is to contrast with black, whereas that of red is to target rods; thus, the function of red is decoupled from that of black rings unlike white/yellow rings); and (3) why false coralsnakes show more pattern irregularities than coralsnakes and have lost coralsnake color patterns more frequently (greater variation in the strength of ophiophagy as a selective force likely stemming from their more diverse foraging habits). Moreover, the clear predictions proposed by Mouy
[66] are testable and permit a direct protective function to be diagnosed.
One final possibility is the proposition that coralsnake colors do not have a single function but perform many functions; that is, the aforementioned explanations for the functions of coralsnake colors are not necessarily mutually exclusive. We think that this is unparsimonious given that it is improbable that the evolution and maintenance of coralsnake colors are driven by several factors. As defined previously, the function of a trait is the effect that selected for the evolution and maintenance of that trait.
Animal colorations can have many effects, like the red coloration of coralsnakes. However, the function of red coloration is only the effect that selected for red, that contributes to increases in redness, and that maintains redness despite selection against it. Although red coloration has several effects, it is unlikely to have several functions; red coloration is a relatively unusual trait, and it is not parsimonious to expect that several effects select for increases in redness. Rather than casting the problem of determining the function of coralsnake color patterns as undecipherably complex because they have many functions, we suggest that biologists focus on identifying the main effect selecting for coralsnake color patterns.
4.4 Conclusion
Aposematism and mimicry are highly complex solutions to the simple problem of avoiding being eaten. The solutions that organisms are prone to evolve in response to this problem in nature tend to be fairly simple (e.g., mammalian herbivores evolve faster flight capabilities to outrun their carnivorous predators, snakes evolve increased venom toxicity to incapacitate their prey more rapidly and prevent counterattacks). The same is likely to be the case for snakes with red and black rings. Over 150 years have passed since the possibility of aposematism and mimicry in coralsnakes was first proposed, yet a single clear case of either of these phenomena in coralsnakes and false coralsnakes is lacking.
Our own field observations and other published observations indicate that coralsnakes and false coralsnakes do not meet many of the putative characteristics of prototypical aposematic and mimetic species, and the existing evidence for aposematism and mimicry in coralsnakes is far from convincing. Rather than assuming an aposematic function for coral snake color patterns, we encourage more research aimed at exploring simpler, alternative solutions with clear testable predictions for the defensive function of coral snake color patterns.
Appendix: Material and Methods
Studies were obtained via weekly searches from January 2021 to March 2024. We performed the searches as follows:
- Weekly searches of Google scholar
- Weekly searches of Scielo;
- Weekly searches of Scopus.
Searches were conducted using the following keywords in different combinations using the Boolean connectors AND OR: snake venoms,
Micrurus,
Leptomicrurus,
Micruroides, transcriptome, proteome, evolution.
Genetic data were retrieved from Genbank for four mitochondrial markers (12s, 16s, cyt b and ND4; Table S1) and one nuclear marker (C-mos), for 44 species of
Micrurus (one specimen per species), plus
Micruroides euryxanthus (1 specimen), and with
Ophiophagus hannah and
Sinomicrurus macclellandi as outgroups (Table S1). Sequences for each genetic marker were aligned using MUSCLE algorithm
[68] as implemented in GENEIOUS 9.1.8
[69] with the default parameters.
In IQTree 2.2.0
[70], we calculated simultaneously the best partition scheme (by gene or codon) and evolutionary models (using ModelFinder,
[71]); the Maximum Likelihood tree, and node support (using 5000 replicates of UltraFast Bootstrap,
[72]). For tree annotation and plotting, we used the R packages ‘ape’
[73], ‘phangorn’
[74] and ‘phytools’
[75] with some posterior edits made using Inkscape 1.3 (www.inkscape.org).
Table S1. Voucher and Genbank accession numbers for the sequences used herein.
| Species |
Voucher |
12S |
16S |
cyt b |
ND4 |
Cmos |
| Micrurus albicinctus |
MPEG19548 |
|
|
|
JF308714 |
|
| Micrurus alleni |
ADM272 |
|
KX660164 |
KX660439 |
KX660567 |
KX660303 |
| Micrurus altirostris |
MCP5035 |
|
|
|
AF228431 |
|
| Micrurus apiatus |
RH52446/M50 |
Z46454 |
Z46484 |
KU754325 |
KU754434 |
|
| Micrurus baliocoryphus |
MZUSP10808 |
|
|
|
AF228433 |
|
| Micrurus bocourti |
MECN2608 |
|
|
|
KP998031 |
|
| Micrurus brasiliensis |
CEPB2301 |
|
|
|
AF228427 |
|
| Micrurus browni |
M223 |
|
|
MG947754 |
MG947664 |
|
| Micrurus carvalhoi |
CEPB2687 |
|
|
|
AF228438 |
|
| Micrurus cf ibiboboca |
IVB1757 |
|
|
|
AF228436 |
|
| Micrurus circinalis |
UWIZM2011.19.7 |
MK534131 |
MK534141 |
MK534160 |
MK534170 |
MK534151 |
| Micrurus corallinus |
USNM253597 |
|
|
|
JF308715 |
|
| Micrurus decoratus |
IVB 2453 |
|
|
|
AF228441 |
|
| Micrurus diastema |
M299 |
|
|
MG947767 |
MG947683 |
|
|
Micrurus dissoleucus |
UTAM673 |
|
|
|
JF308717 |
|
| Micrurus distans |
M301 |
|
|
MG947768 |
MG947685 |
|
| Micrurus diutius |
CAS231787 |
MK534136 |
MK534149 |
MK534168 |
K534178 |
MK534158 |
|
Micrurus dumerilii |
MECN2881 |
|
|
|
KP998035 |
|
| Micrurus elegans |
M48 |
|
|
MG947782 |
MG947710 |
|
| Micrurus ephippifer |
M336 |
|
|
|
MG947697 |
|
|
Micrurus filiformis |
ICN11380 |
MW665121 |
MT862136 |
|
MW662060 |
MW662077 |
| Micrurus frontalis |
CEPB1587 |
|
|
|
AF228425 |
|
| Micrurus fulvius |
Mitohondrion |
GU045453 |
GU045453 |
GU045453 |
GU045453 |
|
|
Micrurus helleri |
AF4455 |
MW665123 |
MW672259 |
MW662069 |
MW662067 |
MW662081 |
| Micrurus ibiboboca |
YPX161 |
YPX161 |
|
|
YPX161 |
|
| Micrurus laticollaris |
UTAM673 |
|
|
|
JF308717 |
|
|
Micrurus lemniscatus |
CAS257629 |
MK534136 |
MK534150 |
MK534169 |
MK534179 |
MK534159 |
| Micrurus mertensi |
CORBIDI14775 |
|
|
|
KP998037 |
|
| Micrurus michoacanensis |
M331 |
|
|
MG947774 |
MG947693 |
|
|
Micrurus mipartitus |
CH5377/USNM348250 |
|
MH140862 |
MH140283 |
EF137406 |
|
| Micrurus mosquitensis |
M12 |
|
|
MG947741 |
MG947650 |
|
|
Micrurus narduccii |
WED54125 |
|
|
EF137412 |
EF137404 |
EF137420 |
| Micrurus nebularis |
M341 |
|
|
MG947778 |
MG947701 |
|
|
Micrurus nigrocinctus |
M120 |
|
|
MG947742 |
MG947651 |
|
|
Micrurus obscurus |
KU222205 |
|
|
|
JF308708 |
|
| Micrurus oliveri |
M632 |
|
|
|
MG947722 |
|
|
Micrurus ornatissimus |
QCAZ2584 |
|
|
|
AF228442 |
|
| Micrurus ortoni |
QCAZ2584 |
|
|
|
AF228442 |
|
|
Micrurus psyches |
ROM28378 |
|
|
|
JF308713 |
|
| Micrurus pyrrhocryptus |
MZUSP10809 |
|
|
|
AF228434 |
|
| Micrurus sp |
CEPB2289 |
|
|
|
AF228439 |
|
| Micrurus stewarti |
CH5408 |
|
MH140870 |
|
KX090915 |
|
|
Micrurus surinamensis |
CEPB8984 |
MN587874 |
MN587874 |
MN587874 |
MN587874 |
MN587874 |
| Micrurus tener |
M200 |
|
|
KU754314 |
KU754411 |
|
| Ophiophagus hannah |
|
NC_011394 |
NC_011394 |
NC_011394 |
NC_011394 |
KX694798 |
| Micruroides euryxanthus |
|
|
|
EF137416 |
EF137408 |
EF137423 |
| Sinomicrurus macclellandi |
|
NC_054255 |
NC_054255 |
NC_054255 |
NC_054255 |
MG653595 |
The map was produced using QGIS 3.34
[76] and edited using Inkscape 1.3 (www.inkscape.org). Samples were names according to their species, notating individuals from different populations with a an abbreviated code at the end of the sample name as follows: AL (Alagoas, Brazil), B (Baurú, Brazil), BR (Brazil), CO (Colombia), EC/EQ (Equador), GO (Goiás, Brazil), J (Jaú, Brazil), PA (Pará, Brazil), RJ (Rio de Janeiro, Brazil), RO (Roraima, Brazil), SP (São Paulo, Brazil), TO (Tocantins, Brazil), UNK (Unknown exact location).
To determine the relationships between different venom toxin classes, we recovered proteomic data of relevant toxins from published works with coral snake species
[9,11,12,77–81]. We transformed the obtained data into percentages to obtain comparable values of the proportion of each toxin class. We then performed an ancestral state reconstruction analysis using the default parameters of the
fastAnc function implemented in the Phytools package
[75] in R software using the phylogeny assembled herein to estimate branch lengths. We built a tree using the
contMap function in Phytools to display continuous variation in toxin proportions for each node using a color gradient.
The left eye of the
Micrurus mipartitus specimen (Panama, Bocas del Toro, Isla San Cristobal; USNM 348250) was scanned using an optical coherence tomography scanner (OQ StrataScope, Lumedica Inc., Durham, NC, USA). A total of 30 B-scans were taken of the eye along both the dorsal–ventral and anterior–posterior axes, and both sets of these 30 B-scans were averaged. The equatorial diameter of the lens was measured by taking the average of measurements from the two averaged B-scans using electronic calipers in Lumedica software. Horizontal spectacle diameter of the left eye was measured using handheld digital calipers with the aid of a stereo microscope.